пролиферация цилиндрического эпителия — 25 рекомендаций на Babyblog.ru
Девочки, привет! Помогите разобраться пожалуйста кто сталкивался…
Все у меня было хорошо, пошла к гинекологу из за того что на Джесе месячные стали скудные слишком. Она взяла стандартные анализы, мазок и цитологию. Пришла за результатами, и они оказались плохие как врач сказала. Надо говорит биопсию делать. Но детально ничего не объяснила( Только напугала и расстроила…
Исследование соскоба с шейки матки цервикального канала
Микроскопическое описание:
Эндоцервикс и Эктоцервикс-скопление клеток цилиндрического эпителия с признаками пролиферации, явления плоскоклеточной метаплазии эпителия, скопление клеток плоского эпителия поверхностного и промежуточного слоев с реактивными изменениями, часть клеток с явлением дискариоза и паракератоза.
Заключение: Возможно, эктопия шейки матки. Рекомендуется исключить HPV.
Уроuенитальные мазки:
Эпителиальные клетки
влагалище: умерено
шейка матки: единичные в поле зрения
эритроциты: не обнаружено
лейкоциты:
влагалище: 20-30
шейка матки: 40-50
флора
влагалище: палочки обильно
шейка матки: палочки умерено
грибы не обнаружено
простейшие (трихоионады) не обнаружено
neisseria gonorroeae не обнаружено
Девченки, кто понимает подскажите по анализам, все настолько плохо?((( Не понимаю я типичнее клетки есть или нет, обычно четко было написано всегда что нет атипичных клеток!
а результат этого анализа я вообще не понимаю. ..Дисплазия что ли? Колпоскопию кстати не делали мне и даже не предлагали, сразу сказала что надо биопсию. ВПЧ есть у меня давно, но проявлений не было никогда. До родов появилась эрозия очень маленькая, в этот раз врач тоже сказала что она есть но очень очень маленькая. Пока назначила просто свечи противовоспалительные. К слову, врачу и клинике я не доверю…просто пошла по ДМС от работы к ним. Они мне в прошлый раз уже ставили внематочную беременность, хотя по сроком просто плодное яйцо еще не опустилось…после этого я очень сомневаюсь в их компетентности. Думаю куда со всем этим теперь идти, может сразу в онко центр? в Герцена…или может в Кулаково?
..Дисплазия что ли? Колпоскопию кстати не делали мне и даже не предлагали, сразу сказала что надо биопсию. ВПЧ есть у меня давно, но проявлений не было никогда. До родов появилась эрозия очень маленькая, в этот раз врач тоже сказала что она есть но очень очень маленькая. Пока назначила просто свечи противовоспалительные. К слову, врачу и клинике я не доверю…просто пошла по ДМС от работы к ним. Они мне в прошлый раз уже ставили внематочную беременность, хотя по сроком просто плодное яйцо еще не опустилось…после этого я очень сомневаюсь в их компетентности. Думаю куда со всем этим теперь идти, может сразу в онко центр? в Герцена…или может в Кулаково?
Частота рака шейки матки в Украине остается высокой и с каждым годом количество заболевших женщин продолжает возрастать
Частота рака шейки матки в Украине остается высокой и с каждым годом количество заболевших женщин продолжает возрастать. Поэтому своевременное выявление, обследование и лечение предраковых поражений, к которым в том числе относятся эрозии шейки матки, имеет большое значение.
О кольпоскопии сказано уже много и каждая женщина, проходившая медицинский осмотр в кабинете врача-гинеколога имела возможность видеть кольпоскоп и изображения шейки матки, которые фиксирует видеосистема. В данной статье хотелось бы остановится на особенностях кольпоскопической картины при некоторых физиологических и патологических состояниях организма женщины
При беременности на шейке матки происходят морфологические изменения, связанные с гормональными изменениями в организме женщины, которые приводят к увеличению в много слойном плоском эпителии количества делящихся клеток, их гиперплазии и изменениям дифференцировки. Шейка матки становится цианотичной (синюшной) и гиперторофированной. При этом наружный зев может выглядеть несколько зияющим, а цилиндрический эпителий цервикального канала незначительно вывернутым наружу, отечным и гиперплазированным.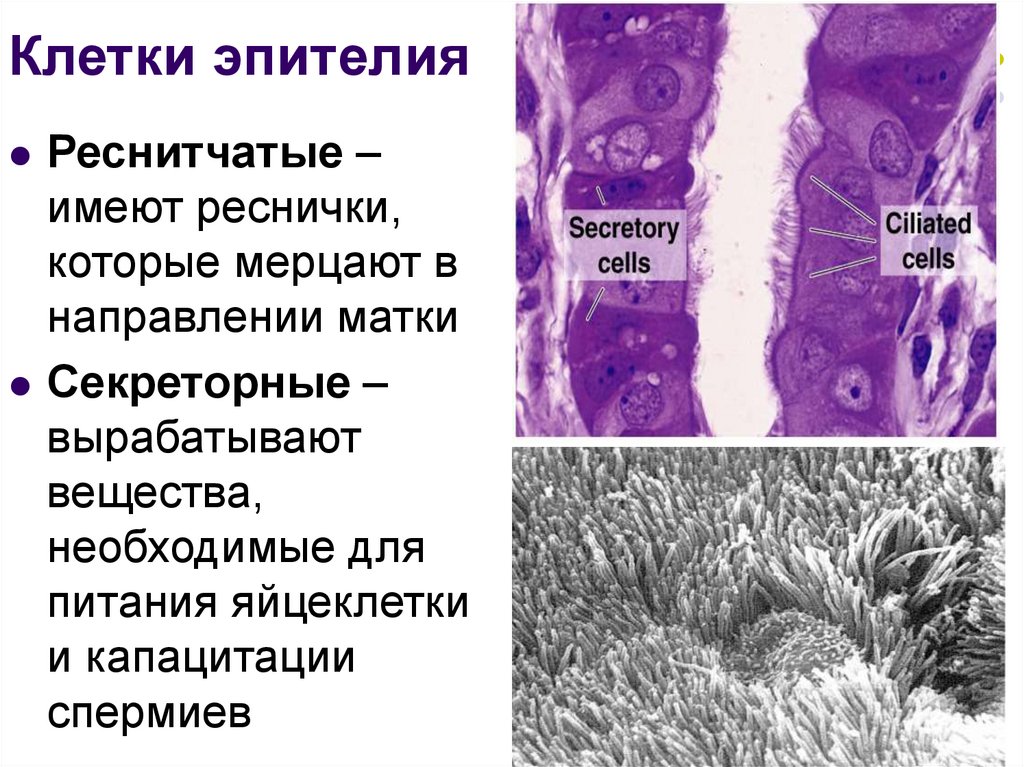
ОСОБЕННОСТИ КОЛЬПОСКОПИИ В ПОСЛЕРОДОВОМ ПЕРИОДЕ
Решение о характере лечения после родов принимается в сроки от 6 недель до 6 месяцев на основании переоценки кольпоскопических результатов обследования.
Особенности кольпоскопии в послеродовом периоде связаны с низким уровнем эстрогенов,что связано с лактацией. Поэтому эпителий шейки матки бледный и при нанесении раствора Люголя окрашивается слабо и неравномерно. Поэтому в сомнительных случаях рекомендуется провести локальный курс эстрогенами с дальнейшим контрольным обследованием.
ОСОБЕННОСТИ КОЛЬПОСКОПИИ В ПОСТМЕНОПАУЗЕ
В результате снижения функции яичников происходят атрофические изменения эпителия шейки матки. Эпителий истончается, под ним видны кровеносные сосуды, из-за недостаточного созревания эпителия меняется биоценоз влагалища и утрачивается естественная защита влагалища, в связи с чем в постменопаузальном периоде часто возникают вторичные воспалительные процессы.
Кольпоскопическая картина определяется истончением и незрелостью эпителия. Сосуды просматриваются через эпителий как множественные красные точки или петли. Такая картина напоминает пунктацию, но при атрофии она наблюдается диффузно на большей части шейки матки. На раствор уксусной кислоты сосуды практически не реагируют, при обработке раствором Люголя эпителий окрашивается неравномерно или не пр. окрашивается совсем. При неясных результатах кольпоскрпии и сомнительных цитологических мазках рекомендуют провести курс лечения эстрогенами с дальнейшим контролем.
Сосуды просматриваются через эпителий как множественные красные точки или петли. Такая картина напоминает пунктацию, но при атрофии она наблюдается диффузно на большей части шейки матки. На раствор уксусной кислоты сосуды практически не реагируют, при обработке раствором Люголя эпителий окрашивается неравномерно или не пр. окрашивается совсем. При неясных результатах кольпоскрпии и сомнительных цитологических мазках рекомендуют провести курс лечения эстрогенами с дальнейшим контролем.
ОСОБЕННОСТИ КОЛЬПОСКОПИИ ПРИ КОНТРАЦЕПЦИИ И ГОРМОНАЛЬНОЙ ТЕРАПИИ
Экзогенные эстрогены стимулируют пролиферацию эпителия шейки матки, в связи с чем появляется паракератоз ( утолщение эпителия) и усиливают процессы репарации и быстрое заживление мелких травм.
При кольпоскопии наблюдается гипертрофия эпителия с повышением продукции слизи. Сосочки цилиндрического эпителия удлиняются и гипертрофируются, иногда с полиповидными складками и увеличенной васкуляризацией.
ОСОБЕННОСТИ КОЛЬПОСКОПИИ ПРИ ВОСПАЛЕНИИ
Характер воспаления зависит от возбудителя и клинически может быть диффузным или локальным, острым или хроническим.
Учитывая, что вирус папилломы человека обладает высоким сродством к эпителиальным тканям и доказано существование высоко- и низкоонкогенных типов вируса, способных вызывать рак шейки матки особое значение приобретает диагностика и выбор тактики лечения при обнаружении ВПЧ. Однако в большинстве случаев кольпоскопические признаки являются неспецифичными. Выделяют экзо фитные формы, которые возвышаются над поверхностью эпителия шейки матки и плоские или субклинические, которые кольпоскопически определяются нормальной и атипической зоной трансформации, ацетон елями участками, не возвышающимися над поверхностью, мозаика, пунктация, негативные участки, атипические сосуды.
Однако в большинстве случаев кольпоскопические признаки являются неспецифичными. Выделяют экзо фитные формы, которые возвышаются над поверхностью эпителия шейки матки и плоские или субклинические, которые кольпоскопически определяются нормальной и атипической зоной трансформации, ацетон елями участками, не возвышающимися над поверхностью, мозаика, пунктация, негативные участки, атипические сосуды.
ОСОБЕННОСТИ КОЛЬПОСКОПИИ ПРИ ЭКЗОФИТНЫХ КОНДИЛОМАХ
Кондиломы представляют собой разрастания соединительной ткани с сосудами внутри, покрытие плоским эпителием, имеют тонкую ножку или широкое основание. Кольпоскопически типичные кондиломы имеют вид пальцеобразных выпячиваний с наличием петли сосуда в каждом из них и выглядят как розовые или сероватые возвышения, расположенные очагово или диффузно. На ранних стадиях кондиломы следует дифференцировать с гиперплазированным цилиндрическим эпителием. При пробе с уксусной кислотой, в отличие от цилиндрического эпителия, кондилома резко белеет, реагируя на кислоту.
Материал подготовил врач-гинеколог Ладная Татьяна Павловна
Мазок на онкоцитологию, пролиферация цилиндрического эпителия — Вопрос гинекологу
Если вы не нашли нужной информации среди ответов на этот вопрос, или же ваша проблема немного отличается от представленной, попробуйте задать дополнительный вопрос врачу на этой же странице, если он будет по теме основного вопроса. Вы также можете задать новый вопрос, и через некоторое время наши врачи на него ответят. Это бесплатно. Также можете поискать нужную информацию в похожих вопросах на этой странице или через страницу поиска по сайту. Мы будем очень благодарны, если Вы порекомендуете нас своим друзьям в социальных сетях.Медпортал 03online.com осуществляет медконсультации в режиме переписки с врачами на сайте. Здесь вы получаете ответы от реальных практикующих специалистов в своей области. В настоящий момент на сайте можно получить консультацию по 71 направлению: специалиста COVID-19, аллерголога, анестезиолога-реаниматолога, венеролога, гастроэнтеролога, гематолога, генетика, гепатолога, гериатра, гинеколога, гинеколога-эндокринолога, гомеопата, дерматолога, детского гастроэнтеролога, детского гинеколога, детского дерматолога, детского инфекциониста, детского кардиолога, детского лора, детского невролога, детского нефролога, детского офтальмолога, детского психолога, детского пульмонолога, детского ревматолога, детского уролога, детского хирурга, детского эндокринолога, дефектолога, диетолога, иммунолога, инфекциониста, кардиолога, клинического психолога, косметолога, логопеда, лора, маммолога, медицинского юриста, нарколога, невропатолога, нейрохирурга, неонатолога, нефролога, нутрициолога, онколога, онкоуролога, ортопеда-травматолога, офтальмолога, паразитолога, педиатра, пластического хирурга, проктолога, психиатра, психолога, пульмонолога, ревматолога, рентгенолога, репродуктолога, сексолога-андролога, стоматолога, трихолога, уролога, фармацевта, физиотерапевта, фитотерапевта, флеболога, фтизиатра, хирурга, эндокринолога.
В настоящий момент на сайте можно получить консультацию по 71 направлению: специалиста COVID-19, аллерголога, анестезиолога-реаниматолога, венеролога, гастроэнтеролога, гематолога, генетика, гепатолога, гериатра, гинеколога, гинеколога-эндокринолога, гомеопата, дерматолога, детского гастроэнтеролога, детского гинеколога, детского дерматолога, детского инфекциониста, детского кардиолога, детского лора, детского невролога, детского нефролога, детского офтальмолога, детского психолога, детского пульмонолога, детского ревматолога, детского уролога, детского хирурга, детского эндокринолога, дефектолога, диетолога, иммунолога, инфекциониста, кардиолога, клинического психолога, косметолога, логопеда, лора, маммолога, медицинского юриста, нарколога, невропатолога, нейрохирурга, неонатолога, нефролога, нутрициолога, онколога, онкоуролога, ортопеда-травматолога, офтальмолога, паразитолога, педиатра, пластического хирурга, проктолога, психиатра, психолога, пульмонолога, ревматолога, рентгенолога, репродуктолога, сексолога-андролога, стоматолога, трихолога, уролога, фармацевта, физиотерапевта, фитотерапевта, флеболога, фтизиатра, хирурга, эндокринолога.
Мы отвечаем на 97.48% вопросов.
Оставайтесь с нами и будьте здоровы!
Столбчатый эпителий — обзор
B Стволовые клетки кишечника и их регулирование ниши
Собственная пластинка и верхний столбчатый эпителий тонкой и толстой кишки усеяны инвагинациями, называемыми криптами. Крипты окружены многочисленными ворсинками, которые выступают в просвет кишечника, создавая большую площадь для абсорбции. Эпителий тонкой кишки поглощает питательные вещества и электролиты и содержит четыре дифференцированных типа клеток: клетку Панета, которая секретирует антимикробные агенты, энтероциты, бокаловидные клетки и энтероэндокринные клетки (Barker et al., 2010а, б; Ван дер Флиер и Клеверс, 2008 г.). Клетки Панета располагаются на дне крипты, тогда как другие клетки занимают поверхность ворсинок. Эпителий толстой кишки содержит два основных дифференцированных типа клеток: абсорбирующие колоноциты и бокаловидные клетки, продуцирующие слизь (Gregorioff and Clevers, 2005). Эпителий толстой кишки реабсорбирует воду и некоторые витамины и поддерживает водный баланс.
Эпителий толстой кишки реабсорбирует воду и некоторые витамины и поддерживает водный баланс.
Дифференцированные типы клеток кишечного эпителия переворачиваются и регенерируются кишечными стволовыми клетками (ISC), находящимися на дне крипт (Brittan and Wright, 2004; Casali and Batile, 2009; Barker et al., 2010а, б). Стволовые клетки крипт также регенерируют эпителий после химического или лучевого поражения (Brittan and Wright, 2004). Исследования импульсного мечения с 3 H-тимидином (рис. 5.2) ясно продемонстрировали, что сначала клетки в нижней части метки крипты, а меченые клетки обнаруживаются прогрессивно дальше по стенке крипты в более позднее время (Potten, 1998). Точное расположение ISC в криптах было неопределенным до тех пор, пока индуцируемая генетическая маркировка не идентифицировала Lgr4 / 5 как криптоспецифические белки в тонком кишечнике и толстой кишке (Barker et al., 2010а, б; Quyn et al., 2010). Ячейки Lgr4 + маркируют все ячейки крипты (ячейки 1–7), тогда как ячейки Lgr5 + маркируют фактические ISC в основании крипты (рис. 5.3). Стволовые клетки Lgr5 + называются столбчатыми клетками с основанием крипт (CBC) и первоначально были идентифицированы Cheng и LeBlond (1974) с использованием метки 3 H-T и электронной микроскопии. Они вставлены между клетками Панета до положения 3 и фактически являются источником клеток Панета (Sato et al., 2011). Положение +4 представляет собой популяцию, промежуточную между клетками CBC и временными амплифицирующими клетками, которые дифференцируются в функциональные клетки ворсинок (Fuchs, 2009; Barker et al., 2010a, b). Клетки +4 обогащены экспрессией гена Bmi , члена семейства polycomb, кодирующего белки, участвующие в регуляции экспрессии генов (Sangiorgi and Capecchi, 2008). И CBC, и +4 клетки также экспрессируют Musashi, а также другие маркеры (Nishimura et al., 2003; Barker et al., 2010а, б). Какое значение могут иметь различия между клетками CBC и клетками +4, неизвестно. Стволовые клетки кишечника экспрессируют фактор транскрипции Tcf-4, который подавляет транскрипцию в сочетании с TLE-1 или CREB-связывающим белком (CBP), поддерживая их в состоянии покоя (Brittan and Wright, 2004).
5.3). Стволовые клетки Lgr5 + называются столбчатыми клетками с основанием крипт (CBC) и первоначально были идентифицированы Cheng и LeBlond (1974) с использованием метки 3 H-T и электронной микроскопии. Они вставлены между клетками Панета до положения 3 и фактически являются источником клеток Панета (Sato et al., 2011). Положение +4 представляет собой популяцию, промежуточную между клетками CBC и временными амплифицирующими клетками, которые дифференцируются в функциональные клетки ворсинок (Fuchs, 2009; Barker et al., 2010a, b). Клетки +4 обогащены экспрессией гена Bmi , члена семейства polycomb, кодирующего белки, участвующие в регуляции экспрессии генов (Sangiorgi and Capecchi, 2008). И CBC, и +4 клетки также экспрессируют Musashi, а также другие маркеры (Nishimura et al., 2003; Barker et al., 2010а, б). Какое значение могут иметь различия между клетками CBC и клетками +4, неизвестно. Стволовые клетки кишечника экспрессируют фактор транскрипции Tcf-4, который подавляет транскрипцию в сочетании с TLE-1 или CREB-связывающим белком (CBP), поддерживая их в состоянии покоя (Brittan and Wright, 2004).
Рисунок 5.2. Слева , схема крипты и ворсинки эпителия слизистой оболочки тонкой кишки. Цифры в склепе указывают положение ячейки.Стрелки указывают, что клетки крипты питают несколько ворсинок. Справа , авторадиограммы 3 H-тимидин меченных крипт / ворсинок через (а) 40 мин, (б) 24 ч, (в) 48 ч, (г) 72 ч. Очевидно движение меченых клеток из глубины крипты (а) к верхушке ворсинки (г).
Воспроизведено с разрешения Potten, Стволовые клетки желудочно-кишечного эпителия: числа, характеристики и гибель. Фил Транс Р. Соц Лондон B 353: 821–830. Авторское право 1998 г., Королевское общество. Рисунок 5.3. Диаграмма, иллюстрирующая обновление эпителия тонкого кишечника кишечными стволовыми клетками.( A ) Общая архитектура крипты, указывающая положения зоны стволовых клеток, промежуточного положения +4 и компартмента транзитного усиления (ТА). ( B ) Получение функциональных эпителиальных клеток кишечника из стволовых клеток Lgr5 + . ( C ) Маркеры для стволовых клеток на основе крипт (CBC) и +4 клеток. ( D ) Клетки Панета обеспечивают важную нишевую среду для стволовых клеток CBC.
( C ) Маркеры для стволовых клеток на основе крипт (CBC) и +4 клеток. ( D ) Клетки Панета обеспечивают важную нишевую среду для стволовых клеток CBC.
Клетки выше положения 7 в крипте ориентируют свои митотические веретена параллельно просвету и подвергаются планарному делению (Caldwel et al., 2007). Используя многофотонную микроскопию, которая позволяет визуализировать всю толщину крипты мыши, Quyn et al. (2010) обнаружили, что клетки в зоне 1-7 ориентируют свои митотические веретена перпендикулярно просвету кишечника, что также характерно для асимметричного деления мышечных сателлитных клеток (см. Главу 6). Помечая ДНК этих клеток с помощью EdU, они показали, что EdU-положительная ДНК сохраняется через несколько делений и локализуется на стороне базальной мембраны эпителия крипты. Это наблюдение соответствует гипотезе «бессмертной цепи» (Cairns, 1975), которая предполагает, что стволовые клетки сохраняют исходную матричную ДНК вместо реплицированной ДНК, чтобы избежать мутаций, вызванных ошибками репликации (см. Комментарий Yamashita, 2010). Фактически, эта базальная сегрегация бессмертной цепи была отменена у мутантов мышей Apc , которые связаны с раком толстой кишки.
Это наблюдение соответствует гипотезе «бессмертной цепи» (Cairns, 1975), которая предполагает, что стволовые клетки сохраняют исходную матричную ДНК вместо реплицированной ДНК, чтобы избежать мутаций, вызванных ошибками репликации (см. Комментарий Yamashita, 2010). Фактически, эта базальная сегрегация бессмертной цепи была отменена у мутантов мышей Apc , которые связаны с раком толстой кишки.
Одиночные клетки CBC, экспрессирующие Lrg5, при культивировании in vitro могут дать начало органоиду (структура ворсинок-крипта), состоящему из крипты с клетками CBC и Панета, а также транзитной зоны амплификации и различных типов дифференцированных ворсинчатые клетки (Sato et al., 2009). Однако органоиды образуются с гораздо большей частотой, если клетки Lgr5 + смешать с клетками Панета. Клетки Панета экспрессируют высокие уровни Wnt3 и Notch лиганда Dll4, а также Egf и Tgf-α (Sato et al., 2011). Поддержание клеток CBC критически зависит от сигналов Notch и, следовательно, от прямого контакта с клетками Панета (Van Es et al. , 2005a, b, 2010; Fre et al., 2005).
, 2005a, b, 2010; Fre et al., 2005).
Wnts способствуют пролиферации клеток CBC. Условная делеция клеток Lgr4 / 5 + у мышей приводит к потере их CBC (De Lau et al., 2011; Кармон и др., 2011). Lgr4 / 5, которые изначально были сиротскими рецепторами, как было показано, связываются с Frz и LRP с образованием рецепторного комплекса для Wnts, который способствует пролиферации CBC. Кроме того, этот комплекс реагирует на факторы роста R-спондина, секретируемые клетками крипт, для дальнейшего усиления пролиферации CBC. Добавление экзогенного Wnt3A к культивируемым органоидам заставляет их трансформироваться в округлые кисты, лишенные каких-либо дифференцированных типов клеток, а ингибирование секреции Wnt небольшой молекулой IWP1 отменяет пролиферацию органоидных клеток.В генетических моделях мышей для потери клеток Панета количество клеток CBC одновременно снижается (Sato et al., 2011). Фактически, сигнатурный ген, экспрессируемый клетками CBC, является геном фактора транскрипции мишени Wnt Achaete scute-like 2 (Ascl2) (Van der Flier et al.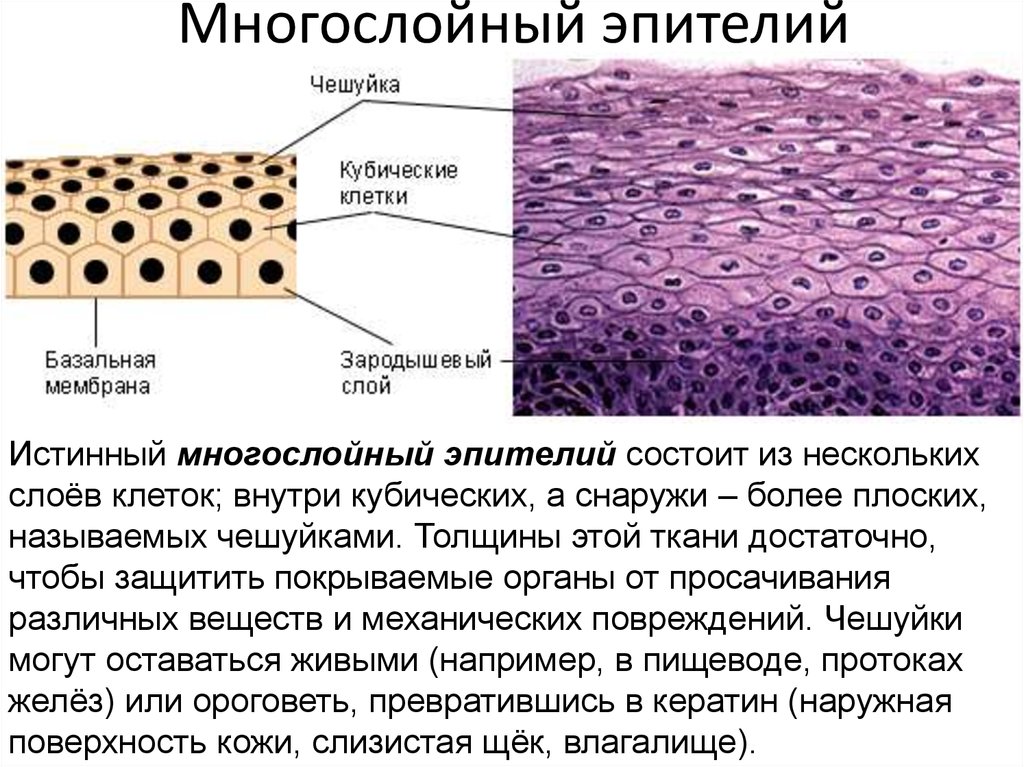 , 2009). Индуцированная делеция гена Ascl2 в тонком кишечнике взрослой мыши приводит к полной потере стволовых клеток Lgr5 + в течение нескольких дней, в то время как трансгенная экспрессия Ascl2 в эпителии кишечника приводит к гиперплазии крипт, включая индукцию эктопические крипты на ворсинах.Эти данные убедительно подтверждают, что клетки Панета используют передачу сигналов Wnt и Notch для контроля количества и судьбы клеток CBC посредством генов-мишеней, таких как Ascl2 .
, 2009). Индуцированная делеция гена Ascl2 в тонком кишечнике взрослой мыши приводит к полной потере стволовых клеток Lgr5 + в течение нескольких дней, в то время как трансгенная экспрессия Ascl2 в эпителии кишечника приводит к гиперплазии крипт, включая индукцию эктопические крипты на ворсинах.Эти данные убедительно подтверждают, что клетки Панета используют передачу сигналов Wnt и Notch для контроля количества и судьбы клеток CBC посредством генов-мишеней, таких как Ascl2 .
Область крипты +4 уступает место большей области транзитных амплифицирующих клеток, которые перемещаются вверх и на ворсинки и участвуют в дифференцировке энтероцитов, бокаловидных клеток и энтероэндокринных клеток. Транзитные усиливающие клетки имеют потенциал до шести делений каждая. Каждая крипта имеет выход 200–300 дифференцированных клеток в день.Количество ISCs в нише крипт строго регулируется апоптозом (Potten et al., 1997). Световая и электронная микроскопия и TUNEL-анализ показали, что апоптоз происходит в 5–10% стволовых клеток в крипте.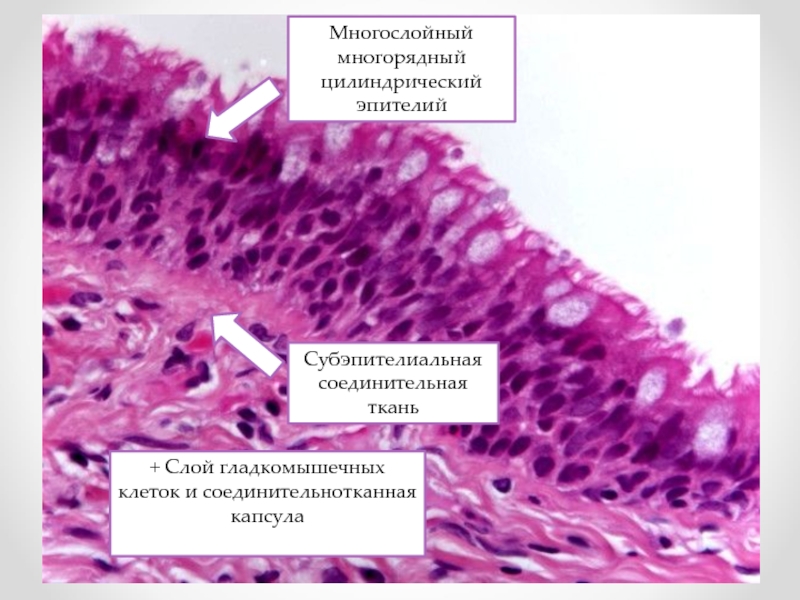
Физиология, эпителизация Статья
Введение
Эпителий состоит из слоев клеток, образующих защитный внешний слой кожи и слизистую оболочку желудочно-кишечного тракта (ЖКТ), мочевыводящих, репродуктивных и дыхательных путей.Этот внешний барьер постоянно поддерживается новыми эпителиальными клетками посредством процесса, называемого эпителизацией. Клинически эпителизация имеет особое значение при заживлении ран. [1]
Проблемы, вызывающие озабоченность
Эпителиальные ткани находятся в коже, желудочно-кишечном тракте, мочевых, репродуктивных и дыхательных путях, где они служат барьером для защиты организма от патогенов и служат для поддержания гомеостаза.Эпителий происходит из каждого из трех слоев эмбриона: эктодермы, энтодермы и мезодермы. [2]
- Форма и расположение эпителиальных клеток различаются в зависимости от расположения и функции тела. Эпителий можно разделить на простые, многослойные, кубовидные и столбчатые эпителиальные клетки.
- Эпителий сохраняет свою прочность благодаря плотным контактам и якорным контактам. Он может передавать молекулы из одной клетки в другую через щелевые соединения.
- Процесс эпителизации жизненно важен для обновления кожи и заживления ран.Эпителиальный слой состоит в основном из кератиноцитов, которые размножаются в базальном слое, дифференцируются, поднимаясь через шиповатый и зернистый слой, а затем теряют ядро и сплющиваются, становясь внешним слоем кожи, известным как роговой слой [1].
- Нарушение процесса эпителизации кожи может вызвать замедленное заживление ран. [1]
Сотовая связь
Существуют общие функциональные и структурные характеристики для всех типов эпителия.Апикальная поверхность эпителиальной клетки обнажается, что позволяет выделять слизь и химические вещества в просвет или открытое пространство. Поверхность эпителиальных клеток, выстилающих структуру тела или орган, называется базальной поверхностью, которая прикрепляется к базальной пластинке, смеси коллагена и гликопротеинов. Базальная пластинка прикрепляет эпителиальную клетку к ретикулярной пластине и образует базальную мембрану. Между соседними мембранами эпителиальных клеток образуются соединения, называемые межклеточными соединениями, обеспечивающие прочность и целостность эпителиального барьера.
Развитие
Эпителиальные клетки происходят из каждого из 3 эмбриональных слоев. Эктодерма развивается в эпителиальную выстилку кожи, носа, рта и заднего прохода. Эндодерма образует эпителий, выстилающий дыхательные пути и большую часть пищеварительной системы. Мезодерма образует эндотелий, который состоит из эпителиальных клеток, выстилающих сосуды и лимфатические сосуды.
Вовлеченные системы органов
Эпителиальные ткани состоят из различных форм и количества слоев, которые способствуют их функции.Эпителий различается в зависимости от анатомического расположения. Плоскоклеточные клетки бывают тонкими и плоскими, кубовидные — квадратными, столбчатыми и прямоугольными. Простой эпителий состоит из одного слоя, в то время как многослойный эпителий состоит из более чем одного слоя уложенных друг на друга эпителиальных клеток [2].
Простой эпителий (однослойных эпителиальных клеток)
Плоский простой эпителий
- Тонкий, плоский, горизонтальный, эллиптический, что позволяет ему быстро проходить через химические вещества.
- Он составляет эндотелий, выстилающий лимфатическую и сердечно-сосудистую системы, альвеолы легких, почечные канальцы и слизистую оболочку капилляров, а также мезотелий, выстилающий серозную поверхность полостей тела и внутренних органов.
Простой кубовидный эпителий
- Коробчатая клетка с круглым ядром; активно впитывает и выделяет
- Обнаружено в канальцах почек и протоках желез
Простой столбчатый эпителий
- Прямоугольной формы с удлиненными ядрами на базальном конце клеток, поглощает и секретирует молекулы.
- Обнаружен в пищеварительном тракте и женских половых путях.
- Ресничные простые столбчатые эпителиальные клетки обнаруживаются в дыхательных путях и фаллопиевых трубах, чтобы перемещать материал по путям.
Псевдостратифицированный столбчатый эпителий
- Он имеет неправильную форму с ядрами в разных местах клетки, и кажется многослойным, но все же представляет собой один слой эпителиальных клеток, все прикрепленные к базальной пластинке.
Многослойный эпителий (клетки, уложенные слоями; защищает от трения и физических сил)
Многослойный плоский эпителий
- Наиболее распространенный тип многослойного эпителия
- Базальный слой кубовидный или столбчатый, а апикальный — плоский
- Верхний слой может быть мертвыми клетками, заполненными кератином
- Обнаружено в коже (ороговевшая) и во рту (не ороговевшая) [2]
Многослойный кубовидный и столбчатый эпителий
- Необычный
- Обнаружено в протоках и железах
Переходный эпителий
- Изменение формы
- Только мочевой пузырь и мочеточники, так как наполнение мочевого пузыря изменяется от кубовидного до плоского.
Железистый эпителий
- Влияет на работу эндокринных и экзокринных желез
- Пот, слюна, грудное молоко
- Тимус, кора надпочечников, гонады, передняя доля гипофиза
- Железы внутренней секреции; высвобождает гормоны непосредственно в ткани
- Экзокринные железы, выделения через проток
Функция
Эпителий служит важным барьером, защищающим внутренние органы от внешних угроз, поддерживая адекватный уровень жидкости в организме и контролируя проницаемость веществ через эпителиальный барьер.[1] Эпителиальные клетки бессосудистые, и питательные вещества проходят через барьер путем абсорбции или диффузии. Они могут очень быстро размножаться, а по мере отмирания клеток они могут отслаиваться в результате процесса, называемого отшелушиванием. В пищеварительном тракте эпителий выделяет пищеварительные ферменты. В дыхательных путях эпителиальные клетки выделяют слизь, которая улавливает микроорганизмы и мусор. Некоторые эпителиальные клетки имеют реснички, микроскопические расширения апикальной мембраны, которые помогают улавливать частицы и перемещать жидкость.В головном мозге реснички помогают циркулировать спинномозговой жидкости (CSF), а в дыхательных путях реснички помогают улавливать патогены в слизистой оболочке. Эпителиальные ткани также могут образовывать экзокринные железы, которые выделяют жидкости через протоки и эндокринные железы, выделяющие гормоны. Клетки эпителия также образуют внешний слой кожи, защищая тело от трения и механических сил. [3]
Механизм
Различные анатомические связи между эпителиальными клетками допускают различные взаимодействия между клетками: плотные соединения, якорные соединения и щелевые соединения.Плотные соединения разделяют эпителиальные клетки на апикальный и базальный отделы. Якорные соединения находятся на базальных и боковых поверхностях эпителиальных клеток и обеспечивают прочность и гибкость тремя различными способами:
- Десмосомы удерживают соседние клетки вместе с помощью молекул адгезии, таких как кадгерин.
- Гемидесмосомы связывают клетки с внеклеточным матриксом с помощью молекул адгезии, таких как интегрины.
- Адгезивные соединения используют актин вместе с кадгерином или интегринами, чтобы влиять на форму эпителия.
- Щелевые соединения уникальны тем, что они образуют внутриклеточную связь между мембранами клеток для переноса молекул ионов и обеспечения возможности эпителия.
Клиническая значимость
Эпителизация кожи
Процесс эпителизации прежде всего изучался в контексте реэпителизации кожи во время заживления ран. Кожа состоит из 3 основных слоев: эпидермиса, дермы и подкожного слоя.Внешний слой кожи состоит из многослойного плоского эпителия, называемого эпидермисом. Эпидермис может быть далее разбит на внешний ороговевший слой, зернистый слой, остистый слой и базальный слой. [1]
Кератиноциты являются основным компонентом эпидермиса и играют ключевую роль в поддержании внешнего кожного барьера и заживлении ран. Кератиноциты пролиферируют в базальном слое, дифференцируются по мере прохождения через гранулярный слой, теряют ядро и сплющиваются, становясь внешним слоем кожи, известным как роговой слой.Недостаток этого процесса может привести к замедленному заживлению ран. [1]
Кератиноциты пролиферируют в базальном слое и состоят из кератиновых промежуточных филаментов K5 и K14. Когда кератиноциты поднимаются в надбазальные слои, они характерно дифференцируются на K1 и K10. В гранулированном слое липиды и белки продуцируются пластинчатыми гранулами и заполняют щели между кератиноцитами в роговом слое. Полимеры с высоким молекулярным весом образуются путем сшивания ороговевших белков оболочки (лорикрин, инволюкрин, филаггрин).Роговая оболочка образуется в процессе терминальной дифференцировки, когда кератиноцит обезвоживается и превращается в многогранник, называемый терминальным корнеоцитом. Липидный слой заполняет корнеоциты, действуя как «ступка», а корнеоциты — как «кирпичи». Этот липидный слой важен для удержания воды в организме. [1]
Во время дифференцировки кератиноцитов клетки переходят от делящихся к неделящимся по мере того, как они мигрируют на поверхность через гранулярный слой.Этот процесс регулируют три основных пути киназы MAP. Эти пути активируются притоком кальция, эпидермальным фактором роста и фактором некроза опухоли. В процессе также используются изоформы протеинкиназы C. Как только кератиноциты превращают профилаггрин в филаггрин, они претерпевают изменения в конце терминальной дифференцировки и необратимо участвуют в процессе дифференцировки. Дифференциация заканчивается, когда протеолитическая и нуклеолитическая активность разрушает клеточные органеллы и ДНК. Повышенный уровень внутриклеточного кальция формирует ороговевшую оболочку за счет активации трансглутаминазы, которая затем ковалентно связывает структурные белки, такие как лорикрин, инволюкрин и филаггрин.Наконец, присоединяются нерастворимые липиды, и завершается внешний слой эпидермиса. [1]
Эпидермальный барьер поддерживается фактором, связанным с кальмодулином кожи (Scarf), который воспринимает кальций и регулирует функцию белка в кожном барьере. Кератиноциты постоянно обновляются, чтобы поддерживать этот барьер. Способность кожи к обновлению обеспечивается популяцией эпидермальных стволовых клеток (ESC). ESC находятся в трех различных нишах: выпуклости волосяных фолликулов, основании сальных желез и базальном слое межфолликулярного эпидермиса.Каждая ниша ESC пополняет свой отсек; следовательно, глубина раны и повреждение этих структур влияет на способность кожи к реэпителизации. [1]
Реэпителизация при заживлении ран
Существует 3 основных фазы заживления ран: воспалительная фаза, фаза пролиферации и фаза дифференцировки. Важно отметить, что эти этапы в значительной степени частично совпадают. Повреждение ткани запускает воспалительную фазу, в которой преобладает высвобождение медиаторов воспаления, нейтрофилов и макрофагов, которые перемещаются к месту раны, чтобы фагоцитировать бактерии и инициировать пролиферативную фазу.Пролиферативная фаза относится к образованию новой грануляционной ткани, заполняющей рану. За этим следует фаза дифференциации, когда волокна коллагена в ране становятся более организованными, а рана укрепляется. [1]
Начальная стадия заживления ран — это воспалительная стадия, вызванная повреждением тканей. Высвобождаются цитокины и вазоактивные вещества, такие как гистамин и серотонин, которые вызывают увеличение проницаемости сосудов. Протекающие сосуды способствуют привлечению лимфоцитов, нейтрофилов и макрофагов к месту раны.Составляющие крови также просачиваются в эту область и активируют каскад свертывания крови. Эти сгустки крови служат матрицей для прикрепления клеток и их миграции в рану и источником факторов роста для фибробластов и воспалительных клеток. В первые 48 часов преобладает воспалительный инфильтрат с нейтрофилами, очищающими рану, и фагоцитирующими бактериями. [1] Также высвобождаются важные цитокины для заживления ран, которые инициируют пролиферативную фазу, такие как IL-1, IL-6, фактор роста эндотелия сосудов, фактор некроза опухоли и TGF-бета.[1] [3]
Когда воспалительная фаза заживления раны заканчивается, начинается пролиферативная фаза, связанная с образованием грануляционной ткани внутри раны. Начало пролиферативной фазы отмечено пролиферацией фибробластов и эндотелиальных клеток. Фибробласты образуют структурные белки, которые производят коллаген и увеличивают прочность раны. Происходит ангиогенез для обеспечения кислородом и питательными веществами новой ткани, гранулирующей в ране. После того, как ложе грануляционной ткани заложено, может начаться процесс реэпителизации.[1]
В течение нескольких часов после травмы кожи кератиноциты по краям раны, вокруг волосяных фолликулов и окружающих сальных желез активируются моноцитами и нейтрофилами, мигрируя и пролиферируя для повторной эпителизации области клетками кожи. [3] Кератиноциты базальных клеток освобождают свое прикрепление к подлежащей дерме и мигрируют через рану, чтобы покрыть ее поверхность, проходя быстрое митотическое деление, поскольку клетки перемещаются друг через друга в манере «чехарда», продвигаясь листом через рану.[1]
Чтобы мигрировать через рану, кератиноциты должны претерпеть структурную трансформацию в клеточном слое. Клетки удлиняются, сглаживаются и развиваются актиновые нити и псевдоподии, которые служат временными выступами для движения [4]. Затем кератиноциты теряют прикрепление полудесмосом к окружающим клеткам. Интегрины (белки для прикрепления) высвобождаются и перемещаются к актиновым филаментам, которые втягивают кератиноциты в рану. Кератиноциты мигрируют во вновь образованный матрикс раны и прикрепляются к ней.Эти изменения в цитоскелете кератиноцитов позволяют увеличить гибкость клеток и миграцию кератиноцитов и обычно отмечаются изменениями в экспрессии кератина-6 и кератина-16. [2] Активированные кератиноциты и фибробласты в дерме высвобождают факторы роста, такие как фактор роста кератиноцитов и фактор роста гепатоцитов, которые помогают регулировать этот процесс. Кератиноциты продолжают размножаться по краям раны и мигрировать, чтобы покрыть рану, пока не встретятся посередине. Некоторые источники сообщают, что хорошо прилегающие раны могут подвергнуться повторной эпителизации в течение 48 часов; другие сообщают, что процесс эпителизации обычно занимает от 2 до 3 недель.[3] Чем быстрее происходит этот процесс, тем меньше остается рубцов. Толстые струпья на ране могут препятствовать способности кератиноцитов перемещаться по ране и увеличивать время заживления.
Через 2 недели рана имеет только 10% общей прочности. [5] Дифференциальная фаза заживления раны происходит от 2 недель до 1 года и связана с уменьшением макрофагов и фибробластов в ране. Фибробласты откладывают в ране коллаген 1 типа, который создает стабильный матрикс [1]. Коллагеновые волокна становятся более организованными, а прочность раны увеличивается.Через 1 месяц рана составляет 50% от общей прочности раны, а через 1 год достигается максимальная прочность рубца 80%.
Если аспекты процесса заживления ран нарушены, это может привести к келоидным рубцам или задержке заживления ран. Протеазы расщепляют пептидные связи и играют важную роль в ремоделировании тканей при заживлении ран. Если это не регулировать, это может привести к хроническим незаживающим кожным язвам. [1]
Заживление ран: роль иммуно-эпителиальных взаимодействий
Peterson, L.W. & Artis, D. Клетки кишечного эпителия: регуляторы барьерной функции и иммунного гомеостаза. Nat. Rev. Immunol. 14 , 141–153 (2014).
CAS PubMed PubMed Central Google ученый
Gurtner, G.C., Werner, S., Barrandon, Y. & Longaker, M.T. Ремонт и регенерация ран. Nature 453 , 314–321 (2008).
CAS Статья Google ученый
Вонг, Дж.W. и др. . Заживление ран на слизистой оболочке полости рта приводит к уменьшению образования рубцов по сравнению с кожей: данные на модели красной свиньи Дюрок и у людей. Регенерация для восстановления ран. 17 , 717–729 (2009).
PubMed Google ученый
Nusrat, A., Delp, C. & Madara, J.L. Реституция эпителия кишечника. Характеристика модели клеточной культуры и картирование элементов цитоскелета в мигрирующих клетках. J. Clin. Инвестировать. 89 , 1501–1511 (1992).
CAS PubMed PubMed Central Google ученый
Янг, Х.В. и др. . Кооперативная активация PI3K малыми GTPases семейства Ras и Rho. Мол. Ячейка 47 , 281–290 (2012).
CAS PubMed PubMed Central Google ученый
Лотц, М.М., Нусрат, А., Мадара, Дж. Л., Эззелл, Р., Вевер, У.М. И Меркурио, А. Реституция кишечного эпителия. Участие специфических изоформ ламинина и рецепторов интегрина ламинина в закрытии ран трансформированного модельного эпителия. Am. J. Pathol. 150 , 747–760 (1997).
CAS PubMed PubMed Central Google ученый
Yamaguchi, N., Mizutani, T., Kawabata, K. & Haga, H. Клетки-лидеры регулируют коллективную миграцию клеток посредством активации Rac в нисходящей передаче сигналов интегрина бета1 и PI3K. Sci. Отчет 5 , 7656 (2015).
CAS PubMed PubMed Central Google ученый
Рэй, Р.М., Маккормак, С.А., Ковингтон, К., Виар, М.Дж., Чжэн, Ю. и Джонсон, Л.Р. Потребность в полиаминах для миграции кишечных эпителиальных клеток опосредуется через Rac1. J. Biol. Chem. 278 , 13039–13046 (2003).
CAS PubMed Google ученый
Баббин, Б.А. и др. . Активация рецептора формилпептида-1 усиливает реституцию кишечных эпителиальных клеток за счет зависимой от фосфатидилинозитол-3-киназы активации Rac1 и Cdc42. J. Immunol. 179 , 8112–8121 (2007).
CAS PubMed Google ученый
Хопкинс, А.М., Пинеда, А.А., Уинфри, Л.М., Браун, Г.Т., Лаукоттер, М.Г. & Nusrat, A. Организованная миграция эпителиальных клеток требует контроля адгезии и выпячивания через эффекторы киназы Rho. Am. J. Physiol. Гастроинтест. Liver Physiol. 292 , G806 – G817 (2007).
CAS PubMed Google ученый
Лейдон, К., Имаидзуми, М., Бартлетт, Р.С., Ван, С.Ф. И Тибо, S.L. Эпителиальные клетки являются активными участниками заживления ран голосовых складок: модель повреждения на животных in vivo, . PLoS One 9 , e115389 (2014).
PubMed PubMed Central Google ученый
Кох, С. и др. . Dkk-1 ингибирует миграцию кишечных эпителиальных клеток, ослабляя направленную поляризацию клеток переднего края. Мол. Биол. Ячейка 20 , 4816–4825 (2009).
CAS PubMed PubMed Central Google ученый
Crosnier, C., Stamataki, D. & Lewis, J. Организация обновления клеток в кишечнике: стволовые клетки, сигналы и комбинаторный контроль. Nat. Преподобный Жене. 7 , 349–359 (2006).
CAS PubMed Google ученый
van der Flier, L.G. & Clevers, H. Стволовые клетки, самообновление и дифференциация кишечного эпителия. Annu. Rev. Physiol. 71 , 241–260 (2009).
CAS PubMed Google ученый
Eisenhoffer, G.T. и др. . Краудинг вызывает экструзию живых клеток для поддержания количества гомеостатических клеток в эпителии. Nature 484 , 546–549 (2012).
CAS PubMed PubMed Central Google ученый
Миёси, Х., Адзима, Р., Луо, К.Т., Ямагути, Т.П. И Стаппенбек, Т. Wnt5a усиливает передачу сигналов TGF-бета, чтобы способствовать регенерации крипт толстой кишки после повреждения ткани. Наука 338 , 108–113 (2012).
CAS PubMed PubMed Central Google ученый
Шейкер, А.И Рубин Д.С. Стволовые клетки: на шаг ближе к восстановлению кишечника. Nature 485 , 181–182 (2012).
CAS PubMed PubMed Central Google ученый
Сато Т. и др. . Единичные стволовые клетки Lgr5 строят структуры крипта-ворсинки in vitro без мезенхимальной ниши. Nature 459 , 262–265 (2009).
CAS PubMed PubMed Central Google ученый
Финни Д.Г. и Прокоп, Д.Дж. Краткий обзор: мезенхимальные стволовые / мультипотентные стромальные клетки: состояние трансдифференцировки и способы восстановления тканей — современные взгляды. Стволовые клетки 25 , 2896–2902 (2007).
PubMed PubMed Central Google ученый
Стаппенбек, Т.С. И Миёси, Х. Роль стромальных стволовых клеток в регенерации тканей и заживлении ран. Наука 324 , 1666–1669 (2009).
CAS PubMed Google ученый
Hong, S.J. и др. . Местно доставляемые стволовые клетки, полученные из жировой ткани, демонстрируют фенотип активированных фибробластов и усиливают образование грануляционной ткани в кожных ранах. PLoS One 8 , e55640 (2013).
CAS PubMed PubMed Central Google ученый
Грин, К.А. и др. .Профибринолитические эффекты металлопротеиназ при заживлении кожных ран в отсутствие плазминогена. J. Invest. Дерматол. 128 , 2092–2101 (2008).
CAS PubMed Google ученый
Kuhn, K.A., Manieri, N.A., Liu, T.C. И Стаппенбек, Т. IL-6 стимулирует пролиферацию кишечного эпителия и восстановление после травм. PLoS One 9 , e114195 (2014).
PubMed PubMed Central Google ученый
Ранкин, К.Р. и др. . Аннексин А2 регулирует интернализацию интегрина бета1 и миграцию кишечных эпителиальных клеток. J. Biol. Chem. 288 , 15229–15239 (2013).
CAS PubMed PubMed Central Google ученый
Алам, А. и др. . Редокс-сигнализация регулирует комменсальный гомеостаз и восстановление слизистой оболочки и требует рецептора формилпептида 1. Mucosal Immunol. 7 , 645–655 (2014).
CAS PubMed Google ученый
Swanson, P.A. II и др. . Кишечные комменсальные бактерии усиливают восстановление эпителия за счет инактивации фосфатаз киназы очаговой адгезии, опосредованной реактивными формами кислорода. Proc. Natl. Акад. Sci. США 108 , 8803–8808 (2011).
CAS PubMed Google ученый
Леони, Г. и др. . Аннексин A1, рецептор формилпептида и NOX1 управляют восстановлением эпителия. J. Clin. Инвестировать. 123 , 443–454 (2013).
CAS PubMed Google ученый
Хант, Т.К., Зедерфельдт, Б. и Голдстик, Т.К. Кислород и исцеление. Am. J. Surg. 118 , 521–525 (1969).
CAS PubMed Google ученый
Вентворт, К.К., Алам, А., Джонс, Р.М., Нусрат, А., Нейш, А.С. Кишечные комменсальные бактерии индуцируют передачу сигналов киназного пути, регулируемого внеклеточными сигналами, посредством зависимой от формилпептидного рецептора редокс-модуляции двойной специфической фосфатазы 3. J. Biol. Chem. 286 , 38448–38455 (2011).
CAS PubMed PubMed Central Google ученый
Чен, К. и др. . Формилпептидный рецептор-2 способствует гомеостазу эпителия толстой кишки, воспалению и онкогенезу. J. Clin. Инвестировать. 123 , 1694–1704 (2013).
CAS PubMed PubMed Central Google ученый
Niethammer, P., Grabher, C., Look, A.T. И Митчисон, Т. Градиент перекиси водорода в масштабе ткани обеспечивает быстрое обнаружение ран у рыбок данио. Nature 459 , 996–999 (2009).
CAS PubMed PubMed Central Google ученый
Ан, Г.О. и др. . Активация транскрипции индуцируемого гипоксией фактора-1 (HIF-1) в миелоидных клетках способствует ангиогенезу через VEGF и S100A8. Proc. Natl. Акад. Sci. США 111 , 2698–2703 (2014).
CAS PubMed Google ученый
Campbell, E.L. и др. . Переносимые нейтрофилы формируют микросреду слизистой оболочки за счет локального дефицита кислорода, что влияет на разрешение воспаления. Иммунитет 40 , 66–77 (2014).
CAS PubMed PubMed Central Google ученый
Louis, N.A., Hamilton, K.E., Kong, T. & Colgan, S.P. HIF-зависимая индукция апикального CD55 координирует эпителиальный клиренс нейтрофилов. FASEB J. 19 , 950–959 (2005).
CAS PubMed PubMed Central Google ученый
Фланниган, К.Л. и др. . Эффекты пролонгирования сероводорода во время колита опосредуются индуцируемым гипоксией фактором-1альфа. FASEB J. 29 , 1531–1602 (2014).
Google ученый
Wallace, J.L., Ferraz, J.G. И Мускара, М. Сероводород: эндогенный медиатор разрешения воспаления и травм. Антиоксид. Редокс-сигнал. 17 , 58–67 (2012).
CAS PubMed PubMed Central Google ученый
Вернер, С.И Гроуз Р. Регулирование заживления ран с помощью факторов роста и цитокинов. Physiol. Ред. 83 , 835–870 (2003).
CAS PubMed PubMed Central Google ученый
Beck, P.L., Rosenberg, I.M., Xavier, R.J., Koh, T., Wong, J.F. & Podolsky, D.K. Трансформирующий фактор роста бета опосредует заживление кишечника и восприимчивость к травмам in vitro, и in vivo, через эпителиальные клетки. Am. J. Pathol. 162 , 597–608 (2003).
CAS PubMed PubMed Central Google ученый
Zindl, C.L. и др. . Нейтрофилы, продуцирующие IL-22, способствуют противомикробной защите и восстановлению целостности эпителия толстой кишки во время колита. Proc. Natl. Акад. Sci. США 110 , 12768–12773 (2013).
CAS PubMed Google ученый
Деннинг, Т.L. & Parkos, C.A. Нейтрофилы используют IL-22 для восстановления порядка в кишечнике. Proc. Natl. Акад. Sci. США 110 , 12509–12510 (2013).
CAS PubMed Google ученый
Пикерт, Г. и др. . STAT3 связывает передачу сигналов IL-22 в эпителиальных клетках кишечника с заживлением ран слизистой оболочки. J. Exp. Med. 206 , 1465–1472 (2009).
CAS PubMed PubMed Central Google ученый
Гриннелл, Ф., Хо, Ч. И Высоцкий, А. Деградация фибронектина и витронектина в хронической раневой жидкости: анализ клеточным блоттингом, иммуноблоттингом и анализами клеточной адгезии. J. Invest. Дерматол. 98 , 410–416 (1992).
CAS PubMed Google ученый
Фуката, М. и др. . Cox-2 регулируется Toll-подобным рецептором-4 (TLR4): роль в пролиферации и апоптозе в кишечнике. Гастроэнтерология 131 , 862–877 (2006).
CAS PubMed PubMed Central Google ученый
Морто, О. и др. . Нарушение защиты слизистой оболочки от острого повреждения толстой кишки у мышей, лишенных циклооксигеназы-1 или циклооксигеназы-2. J. Clin. Инвестировать. 105 , 469–478 (2000).
CAS PubMed PubMed Central Google ученый
Castaneda, C.A. И Гомес, Х.L. Таргетная терапия: комбинация лапатиниба и паклитаксела при HER2-положительном раке молочной железы. Nat. Преподобный Clin. Онкол. 6 , 308–309 (2009).
CAS PubMed Google ученый
Мартин, П. и др. . Заживление ран у мышей без PU.1 — восстановление тканей не зависит от воспалительных клеток. Curr. Биол. 13 , 1122–1128 (2003).
CAS Google ученый
Ламмерманн, Т. и др. . Рой нейтрофилов требует LTB4 и интегринов на сайтах гибели клеток in vivo . Nature 498 , 371–375 (2013).
PubMed Google ученый
Эминг, С.А., Криг, Т. и Дэвидсон, Дж.М. Воспаление при заживлении ран: молекулярные и клеточные механизмы. J. Invest. Дерматол. 127 , 514–525 (2007).
CAS PubMed Google ученый
Демидова-Райс, Т.Н., Хамблин, М. Adv. Уход за кожными ранами 25 , 349–370 (2012).
PubMed PubMed Central Google ученый
Раттер, доктор медицины и др. . Тридцатилетний анализ программы колоноскопического наблюдения за неоплазией при язвенном колите. Гастроэнтерология 130 , 1030–1038 (2006).
PubMed Google ученый
Нуршар С. и Алон Р. Миграция лейкоцитов в воспаленные ткани. Иммунитет 41 , 694–707 (2014).
CAS PubMed PubMed Central Google ученый
Энгельгардт, Э., Токсой, А., Гебелер, М., Дебус, С., Брокер, Э.B. & Gillitzer, R. Хемокины IL-8, GROalpha, MCP-1, IP-10 и Mig последовательно и дифференцированно экспрессируются во время фазоспецифической инфильтрации субпопуляций лейкоцитов при заживлении ран у человека. Am. J. Pathol. 153 , 1849–1860 (1998).
CAS PubMed PubMed Central Google ученый
Sumagin, R. & Sarelius, I.H. Обогащение молекулы-1 межклеточной адгезии вблизи трехклеточных эндотелиальных соединений предпочтительно связано с трансмиграцией лейкоцитов и сигналами для реорганизации этих соединений для обеспечения прохождения лейкоцитов. J. Immunol. 184 , 5242–5252 (2010).
CAS PubMed PubMed Central Google ученый
Schauer, C. и др. . Агрегированные внеклеточные ловушки нейтрофилов ограничивают воспаление за счет разложения цитокинов и хемокинов. Nat. Med. 20 , 511–517 (2014).
CAS PubMed Google ученый
Ву, М.С. и др. . Рецептор для компонента комплемента C3a опосредует защиту от кишечных ишемических-реперфузионных повреждений, ингибируя мобилизацию нейтрофилов. Proc. Natl. Акад. Sci. США 110 , 9439–9444 (2013).
CAS PubMed Google ученый
Sinno, H. et al. . Ускоренное заживление ран при местном применении комплемента C5. Пласт. Реконстр. Surg. 130 , 523–529 (2012).
CAS PubMed Google ученый
Nauseef, W.M. & Borregaard, Н. Нейтрофилы в действии. Nat. Иммунол. 15 , 602–611 (2014).
CAS PubMed Google ученый
Hirsch, T. et al. . Человеческий бета-дефенсин-3 способствует заживлению инфицированных диабетических ран. J. Gene Med. 11 , 220–228 (2009).
CAS PubMed Google ученый
Aarbiou, J. et al. . Дефенсины нейтрофилов усиливают закрытие эпителиальной раны легких и экспрессию гена муцина in vitro . Am. J. Respir. Cell Mol. Биол. 30 , 193–201 (2004).
CAS PubMed Google ученый
Де, Ю. и др. . LL-37, кателицидин, полученный из гранул и эпителиальных клеток нейтрофилов, использует рецептор формилпептида-подобный 1 (FPRL1) в качестве рецептора для хемоаттракции нейтрофилов периферической крови человека, моноцитов и Т-клеток. J. Exp. Med. 192 , 1069–1074 (2000).
Google ученый
Шайхиев Р. и др. . Человеческий эндогенный антибиотик LL-37 стимулирует пролиферацию эпителиальных клеток дыхательных путей и закрытие ран. Am. J. Physiol. Легочная клетка. Мол. Physiol. 289 , L842 – L848 (2005).
CAS PubMed Google ученый
Эшкрофт, Г.С. и др. . Ингибитор секреторной лейкоцитарной протеазы обеспечивает неизбыточные функции, необходимые для нормального заживления ран. Nat. Med. 6 , 1147–1153 (2000).
CAS PubMed Google ученый
Serhan, C.N., Chiang, N. & Van Dyke, T.E. Устранение воспаления: двойные противовоспалительные и способствующие разрешению липидные медиаторы. Nat. Rev. Immunol. 8 , 349–361 (2008).
CAS PubMed PubMed Central Google ученый
Спайт, М. и Серхан, К.Н. Новые липидные медиаторы способствуют разрешению острого воспаления: воздействие аспирина и статинов. Circ. Res. 107 , 1170–1184 (2010).
CAS PubMed PubMed Central Google ученый
Zemans, R.L. и др. . Трансмиграция нейтрофилов запускает восстановление эпителия легких посредством передачи сигналов бета-катенина. Proc. Natl. Акад. Sci. США 108 , 15990–15995 (2011).
CAS PubMed Google ученый
Aoi, Y., Terashima, S., Ogura, M., Nishio, H., Kato, S. & Takeuchi, K. Роли оксида азота (NO) и NO-синтаз в исцелении от декстрансульфата натрия -индуцированный колит у крыс. J. Physiol. Pharmacol. 59 , 315–336 (2008).
CAS PubMed Google ученый
Гинзберг, Х.Х. и др. . Опосредованное нейтрофилами повреждение эпителия во время трансмиграции: роль эластазы. Am. J. Physiol. Гастроинтест. Liver Physiol. 281 , G705 – G717 (2001).
CAS PubMed PubMed Central Google ученый
Таузин, С., Старнес, Т.В., Беккер, Ф. Б., Лам, П. Я. & Huttenlocher, A. Передача сигналов киназ семейства Redox и Src контролирует привлечение лейкоцитов к ране и обратную миграцию нейтрофилов. J. Cell Biol. 207 , 589–598 (2014).
CAS PubMed PubMed Central Google ученый
Xavier, R.J. & Подольский, Д. Раскрытие патогенеза воспалительного заболевания кишечника. Nature 448 , 427–434 (2007).
CAS Google ученый
Fournier, B.M. И Паркос, К.А. Роль нейтрофилов при воспалении кишечника. Mucosal Immunol. 5 , 354–366 (2012).
CAS PubMed PubMed Central Google ученый
Сумагин Р., Робин А.З., Нусрат А. и Паркос К.А. Трансмиграционные нейтрофилы в просвете кишечника задействуют ICAM-1 для регулирования эпителиального барьера и рекрутирования нейтрофилов. Mucosal Immunol. 7 , 905–915 (2014).
CAS PubMed Google ученый
Симпсон Д.М. и Росс, Р. Исследование нейтрофильных лейкоцитов в заживлении ран с использованием антинейтрофильной сыворотки. J. Clin. Инвестировать. 51 , 2009–2023 (1972).
CAS PubMed PubMed Central Google ученый
Souza, C.M. и др. . Регенерации кожной ткани способствуют мезенхимальные стволовые клетки, засеянные наноструктурированной мембраной. Пересадка. Proc. 46 , 1882–1886 (2014).
CAS PubMed Google ученый
Смит П.К., Касерес, М., Мартинес, К., Оярсун, А. и Мартинес, Дж. Заживление десневой раны: существенная реакция, нарушаемая старением? J. Dent. Res. 94 , 395–402 (2014).
PubMed Google ученый
Дови, Дж.В., Хе, Л.К. И ДиПьетро, Л.А. Ускоренное закрытие ран у мышей с истощением нейтрофилов. J. Leukoc. Биол. 73 , 448–455 (2003).
CAS PubMed Google ученый
Нишио, Н., Окава, Ю., Сакураи, Х. и Исобе, К. Истощение нейтрофилов задерживает заживление ран у старых мышей. Возраст (Дордр) 30 , 11–19 (2008).
CAS Google ученый
Le’Negrate, G., Rostagno, P., Auberger, P., Rossi, B. & Hofman, P. Подавление экспрессии каспаз и лиганда Fas и увеличение продолжительности жизни нейтрофилов после трансмиграции через эпителий кишечника. Cell Death Differ. 10 , 153–162 (2003).
CAS PubMed Google ученый
Мюррей П.Дж. и Винн Т.А. Защитные и патогенные функции субпопуляций макрофагов. Nat. Rev. Immunol. 11 , 723–737 (2011).
CAS PubMed PubMed Central Google ученый
Синдрилару, А. и др. . Безудержная провоспалительная популяция макрофагов M1, индуцированная железом, ухудшает заживление ран у людей и мышей. J. Clin. Инвестировать. 121 , 985–997 (2011).
CAS PubMed PubMed Central Google ученый
Мюррей, П.Дж. и др. . Активация и поляризация макрофагов: номенклатура и экспериментальные рекомендации. Иммунитет 41 , 14–20 (2014).
CAS PubMed PubMed Central Google ученый
Мюррей, П.J. Первичный механизм противовоспалительного ответа, регулируемого IL-10, заключается в избирательном ингибировании транскрипции. Proc. Natl. Акад. Sci. США 102 , 8686–8691 (2005).
CAS PubMed Google ученый
Далли, Дж. и др. . Новый 13S, 14S-эпокси-марезин превращается макрофагами человека в марезин 1 (MaR1), ингибирует лейкотриен-А4-гидролазу (LTA4H) и изменяет фенотип макрофагов. FASEB J. 27 , 2573–2583 (2013).
CAS PubMed PubMed Central Google ученый
Hong, S. et al. . Нейропротектин / протектин D1: эндогенный биосинтез и действие на диабетические макрофаги, способствуя заживлению ран и иннервации, нарушенной диабетом. Am. J. Physiol. Cell Physiol. 307 , C1058 – C1067 (2014).
CAS PubMed PubMed Central Google ученый
Шаффер, М.Р., Тантри, У., ван Везеп, Р.А. & Барбул, А. Метаболизм оксида азота в ранах. J. Surg. Res. 71 , 25–31 (1997).
CAS PubMed Google ученый
Stallmeyer, B., Kampfer, H., Kolb, N., Pfeilschifter, J. & Frank, S. Функция оксида азота в заживлении ран: ингибирование индуцируемой синтазы оксида азота серьезно ухудшает реэпителизацию раны. J. Invest. Дерматол. 113 , 1090–1098 (1999).
CAS PubMed Google ученый
Xiao, W., Hong, H., Kawakami, Y., Lowell, C.A. И Каваками Т. Регулирование миелопролиферации и программирование макрофагов M2 у мышей с помощью Lyn / Hck, SHIP и Stat5. J. Clin. Инвестировать. 118 , 924–934 (2008).
CAS PubMed PubMed Central Google ученый
Дигельманн, Р.Ф., Коэн, И. И Каплан, А. Роль макрофагов в заживлении ран: обзор. Пласт. Реконстр. Surg. 68 , 107–113 (1981).
CAS PubMed Google ученый
Neurath, M.F. И Трэвис, С.П. Заживление слизистой оболочки при воспалительных заболеваниях кишечника: систематический обзор. Кишечник 61 , 1619–1635 (2012).
CAS PubMed Google ученый
Нагаока, Т. и др. . Замедленное заживление ран в отсутствие экспрессии молекулы-1 межклеточной адгезии или L-селектина. Am. J. Pathol. 157 , 237–247 (2000).
CAS PubMed PubMed Central Google ученый
Исида, Ю., Гао, Дж. Л. и Мерфи, П.М. Хемокиновый рецептор CX3CR1 опосредует заживление кожных ран, способствуя накоплению и функционированию макрофагов и фибробластов. J. Immunol. 180 , 569–579 (2008).
CAS PubMed Google ученый
Беттенворт, Д. и др. . Трипептид KdPT защищает от воспаления кишечника и поддерживает барьерную функцию кишечника. Am. J. Pathol. 179 , 1230–1242 (2011).
CAS PubMed PubMed Central Google ученый
Кран, М.Дж., Дейли, Дж. М., Ван Хаутт, О., Бранкато, С. К., Генри, В. Л. Jr & Albina, J.E. Переход от моноцитов к макрофагам в стерильной ране мыши. PLoS One 9 , e86660 (2014).
PubMed PubMed Central Google ученый
Винн Т.А., Чавла А. и Поллард Дж. У. Биология макрофагов в развитии, гомеостазе и болезнях. Nature 496 , 445–455 (2013).
CAS PubMed PubMed Central Google ученый
Витте, М.Б. и Барбул А. Общие принципы заживления ран. Surg. Clin. North Am. 77 , 509–528 (1997).
CAS PubMed Google ученый
Pull, S.L., Doherty, J.M., Mills, J.C., Gordon, J.I. И Стаппенбек, Т. Активированные макрофаги являются адаптивным элементом ниши предшественников эпителиального эпителия толстой кишки, необходимого для регенеративных реакций на повреждение. Proc. Natl. Акад. Sci. США 102 , 99–104 (2005).
CAS PubMed Google ученый
Сено, Х., Миёси, Х., Браун, С.Л., Геске, М.Дж., Колонна, М., и Стаппенбек, Т.С. Для эффективного заживления ран слизистой оболочки толстой кишки требуется передача сигналов Trem2. Proc. Natl. Акад. Sci. США 106 , 256–261 (2009).
CAS PubMed Google ученый
Лукас, Т. и др. . Различная роль макрофагов в различных фазах восстановления кожи. J. Immunol. 184 , 3964–3977 (2010).
CAS PubMed Google ученый
Амини-Ник, С. и др. . Адгезия и миграция миелоидных клеток, регулируемые бета-катенином, определяют заживление ран. J. Clin. Инвестировать. 124 , 2599–2610 (2014).
CAS PubMed PubMed Central Google ученый
Нейрат, М.F. Новые мишени для заживления слизистых оболочек и терапии воспалительных заболеваний кишечника. Mucosal Immunol. 7 , 6–19 (2014).
CAS PubMed Google ученый
Далли, Дж. и др. . Проразрешение и тканезащитное действие устойчивых к расщеплению пептидов на основе аннексина A1 опосредуются рецептором формилпептида 2 / рецептором липоксина A4. J. Immunol. 190 , 6478–6487 (2013).
CAS PubMed Google ученый
Drechsler, M. et al. . Аннексин A1 противодействует рекрутированию артериальных миелоидных клеток, вызванному хемокинами. Circ. Res. 116 , 827–835 (2015).
CAS PubMed Google ученый
Леони, Г. и др. . Внеклеточные везикулы и полимерные наночастицы, содержащие аннексин А1, способствуют заживлению эпителиальной раны. J. Clin. Инвестировать. 125 , 1215–1227 (2015).
PubMed PubMed Central Google ученый
Фредман, Г. и др. . Направленные наночастицы, содержащие прорезавляющий пептид Ac2-26, защищают от развитого атеросклероза у мышей с гиперхолестеринемией. Sci. Пер. Med. 7 , 275ра20 (2015).
CAS PubMed PubMed Central Google ученый
Кэш, Дж.L., Bena, S., Headland, S.E., McArthur, S., Brancaleone, V. и Perretti, M. Chemerin15 ингибирует опосредованное нейтрофилами сосудистое воспаление и ишемию-реперфузию миокарда с помощью ChemR23. EMBO Rep. 14 , 999–1007 (2013).
CAS PubMed PubMed Central Google ученый
Serhan, C.N. Липидные медиаторы, способствующие расслоению, являются ведущими для физиологии разрешения. Nature 510 , 92–101 (2014).
CAS PubMed PubMed Central Google ученый
Бакли, К.Д., Гилрой, Д.В. И Серхан, К. Проработка липидных медиаторов и механизмов разрешения острого воспаления. Иммунитет 40 , 315–327 (2014).
CAS PubMed PubMed Central Google ученый
Гронерт, К. Липоксины в глазу и их роль в заживлении ран. Prostaglandins Leukot. Ессент. Жирные кислоты (2005) 73 , 221–229.
CAS PubMed Google ученый
Gronert, K., Maheshwari, N., Khan, N., Hassan, IR, Dunn, M. & Laniado Schwartzman, M. Роль 12/15-липоксигеназного пути мыши в ускорении заживления эпителиальных ран и защита хозяев. J. Biol. Chem. 280 , 15267–15278 (2005).
CAS PubMed Google ученый
Фредман, Г. и др. . Резолвин D1 ограничивает ядерную локализацию 5-липоксигеназы и синтез лейкотриена B4, ингибируя путь киназы, активируемой кальцием. Proc. Natl. Акад. Sci. США 111 , 14530–14535 (2014).
CAS PubMed Google ученый
Далли, Дж., Норлинг, Л.В., Реншоу, Д., Купер, Д., Люнг, К. & Perretti, M. Аннексин 1 опосредует быстрое противовоспалительное действие микрочастиц нейтрофилов. Кровь 112 , 2512–2519 (2008).
CAS PubMed Google ученый
Норлинг, Л.В., Спайт, М., Янг, Р., Флауэр, Р.Дж., Перретти, М. и Серхан, К.Н. Передний край: гуманизированные наноразлагающие лекарства имитируют разрешение воспаления и ускоряют заживление ран. J. Immunol. 186 , 5543–5547 (2011).
CAS PubMed PubMed Central Google ученый
Серхан, К.N. Фаза разрешения воспаления: новые эндогенные противовоспалительные и проресаживающие липидные медиаторы и пути. Annu. Rev. Immunol. 25 , 101–137 (2007).
CAS PubMed Google ученый
Далли, Дж., Чанг, Н. и Серхан, К. Идентификация сульфидо-конъюгированных медиаторов 14-й серии, которые способствуют разрешению инфекции и защите органов. Proc. Natl. Акад. Sci. США 111 , E4753 – E4761 (2014).
CAS PubMed Google ученый
Спайт, М. и др. . Резолвин D2 является мощным регулятором лейкоцитов и контролирует микробный сепсис. Nature 461 , 1287–1291 (2009).
CAS PubMed PubMed Central Google ученый
Sousounis, K., Baddour, J.A. И Цонис, П.А. Старение и регенерация позвоночных. Curr. Верхний. Dev. Биол. 108 , 217–246 (2014).
CAS PubMed Google ученый
Арнардоттир, Х.Х., Далли, Дж., Колас, Р.А., Шинохара, М., Серхан, К.Н. Старение задерживает разрешение острого воспаления у мышей: перепрограммирование реакции хозяина с помощью новых лекарств с наноразрешением. J. Immunol. 193 , 4235–4244 (2014).
CAS PubMed PubMed Central Google ученый
Секи, Х. и др. . Противовоспалительный и проресаживающий медиатор резолвин E1 защищает мышей от бактериальной пневмонии и острого повреждения легких. J. Immunol. 184 , 836–843 (2010).
CAS PubMed Google ученый
Ридер, Ф. и др. . Результаты 2-го научного семинара ECCO (III): основные механизмы заживления кишечника. J. Crohns Colitis 6 , 373–385 (2012).
PubMed Google ученый
Демидова-Райс, Т.Н., Хамблин, М.Р. и Герман, И.М.Острое и нарушенное заживление ран: патофизиология и современные методы доставки лекарств, часть 1: нормальные и хронические раны: биология, причины и подходы к лечению. Adv. Уход за кожными ранами 25 , 304–314 (2012).
PubMed PubMed Central Google ученый
Ридер, Ф., Brenmoehl, J., Leeb, S., Scholmerich, J. & Rogler, G. Заживление ран и фиброз при кишечных заболеваниях. Кишечник 56 , 130–139 (2007).
CAS PubMed PubMed Central Google ученый
Diegelmann, R.F. Избыточные нейтрофилы характерны для хронических пролежней. Регенерация для восстановления ран. 11 , 490–495 (2003).
PubMed Google ученый
Мартин, Дж.М., Зенилман, Дж. М. и Лазарус, Г. С. Молекулярная микробиология: новые измерения кожной биологии и заживления ран. J. Invest. Дерматол. 130 , 38–48 (2010).
CAS PubMed Google ученый
Высоцки, А.Б., Стаяно-Коико, Л. и Гриннелл, Ф. Раневая жидкость хронических язв ног содержит повышенные уровни металлопротеиназ ММР-2 и ММП-9. J. Invest. Дерматол. 101 , 64–68 (1993).
CAS PubMed Google ученый
Эминг, С.А., Мартин, П. и Томич-Каник, М. Восстановление и регенерация ран: механизмы, передача сигналов и трансляция. Sci. Пер. Med. 6 , 265ср6 (2014).
PubMed PubMed Central Google ученый
Ягер, Д.Р., Чжан, Л.Ю., Лян, Х.Х., Дигельманн, Р.Ф. И Коэн, И.Рневые жидкости от пролежней человека содержат повышенные уровни и активность матриксной металлопротеиназы по сравнению с хирургическими раневыми жидкостями. J. Invest. Дерматол. 107 , 743–748 (1996).
CAS PubMed Google ученый
Лауэр, Г. и др. . В хронических ранах повышается экспрессия и протеолиз фактора роста эндотелия сосудов. J. Invest. Дерматол. 115 , 12–18 (2000).
CAS PubMed Google ученый
фон Лампе, Б., Бартель, Б., Коупленд, С.Е., Рикен, Э.О. & Rosewicz, S. Дифференциальная экспрессия матриксных металлопротеиназ и их тканевых ингибиторов в слизистой оболочке толстой кишки пациентов с воспалительным заболеванием кишечника. Кишечник 47 , 63–73 (2000).
CAS PubMed PubMed Central Google ученый
Кастанеда, Ф.Е. и др. . Нацеленная делеция металлопротеиназы 9 ослабляет экспериментальный колит у мышей: центральная роль эпителиального ММП. Гастроэнтерология 129 , 1991–2008 (2005).
CAS PubMed Google ученый
Нава П., Камекура Р. и Нусрат А. Расщепление белков трансмембранного соединения и их роль в регулировании эпителиального гомеостаза. Тканевые барьеры 1 , e24783 (2013).
PubMed PubMed Central Google ученый
Leeb, S.N. и др. . Снижение миграции фибробластов при воспалительном заболевании кишечника: роль медиаторов воспаления и киназы очаговой адгезии. Гастроэнтерология 125 , 1341–1354 (2003).
CAS PubMed Google ученый
Тонг, Кв. и др. . Интерферон-гамма ингибирует миграцию эпителиальных клеток Т84, перенаправляя трансцитоз интегрина бета1 с переднего края миграции. J. Immunol. 175 , 4030–4038 (2005).
CAS PubMed Google ученый
Клеточные изменения и адаптивные реакции
Последнее обновление: 5 августа 2021 г.
Резюме
Клеточная адаптация — это способность клеток реагировать на различные типы стимулов и неблагоприятные изменения окружающей среды. Эти адаптации включают гипертрофию (увеличение отдельных клеток), гиперплазию (увеличение числа клеток), атрофию (уменьшение размера и числа клеток), метаплазию (переход от одного типа эпителия к другому) и дисплазию (нарушение роста клеток).Ткани адаптируются по-разному в зависимости от репликативных характеристик клеток, составляющих ткань. Например, лабильная ткань, такая как кожа, может быстро воспроизводиться и, следовательно, также может регенерироваться после травмы, тогда как постоянная ткань, такая как нервная и сердечная ткань, не может восстанавливаться после травмы. Если клетки не способны адаптироваться к неблагоприятным изменениям окружающей среды, гибель клеток происходит физиологически в форме апоптоза или патологически в форме некроза. В этой статье представлен обзор основных клеточных адаптивных механизмов и их различных последствий в организме человека.
Клеточная адаптация
| Обзор клеточной адаптации | ||
|---|---|---|
| Определение | Формы и примеры | |
| Атрофия |
| |
| Дистрофия |
| |
| Гипертрофия |
| |
| Гиперплазия |
|
|
| Анаплазия | ||
| Метаплазия |
| |
| Дисплазия | ||
| Увеличение | 5 и пролиферация количество ячеек |
|
| Регенерация |
| |
| ||
Повреждение клетки
Стадии
- Ранняя стадия: характеризуется обратимым клеточным набуханием (например, отечная дегенерация)
- Поздняя стадия: характеризуется необратимым повреждением мембраны и гибелью клеток
Причины
Ишемия
-
Описание
- Снижение кровоснабжения, которое не может удовлетворить потребность органа или ткани в кислороде
- Если не исправить, ишемия может привести к гибели клеток из-за набухания клеток (онкоз).
-
Патогенез
- Снижение артериальной перфузии (например, из-за атеросклероза, тромбоэмболии) в солидных органах с единственным (концевым) кровоснабжением (например, почки, сердце) → бледный инфаркт
- Снижение венозного оттока (например, венозная окклюзия, синдром Бадда-Киари, перекрут яичка, перекрут яичника) в тканях с более чем одним кровоснабжением (например, кишечник, легкие, печень, яички) или реперфузия (например, после ангиопластики) → красный инфаркт
- Амортизатор со следующими вариантами:
-
Время толерантности к ишемии: время, по истечении которого ишемия вызывает необратимое повреждение ткани
- Кожа: 12 часов
- Мускулатура: 6–8 часов
- Нервная ткань: 2–4 часа
Повреждение свободными радикалами
Определение
Патофизиология
Возникновение
Кислородная токсичность
[7] [8]Реперфузионное повреждение
Болезни накопления металлов
Химическая смерть /
лекарственная токсичность Смерть клеток — это необратимое повреждение, из-за которого клетки не могут выполнять свои функции.Это приводит либо к апоптозу, либо к некрозу.| Обзор апоптоза и некроза | ||
|---|---|---|
| Характеристики | Апоптоз | Некроз |
| Определение |
|
|
| Патофизиология |
| |
| Микроскопия | ||
Апоптоз
Общая информация
- Определение: запрограммированная гибель клеток (физиологический оборот клеток)
- Этиология
- Характеристики
- Гистопатологические данные
- Апоптоз может инициироваться двумя разными путями: внешним путем (через внешние стимулы) или внутренним путем (через внутренние стимулы).
- Общая последовательность событий: стимул → активация инициаторных каспаз → активация каспаз-палачей → апоптоз
Внешний путь (путь рецептора смерти)
Может быть активирован с помощью 2 механизмов:
Внутренний путь (митохондриальный путь)
- Участвует в ремоделировании тканей (например, во время эмбриогенеза)
- p53 активируется посредством повреждения ДНК (например, гипоксия, химические токсины, радиация) или изъятия регулирующих факторов из популяции пролиферирующих клеток (например.g., IL-2 после завершения иммунологической реакции → апоптоз эффекторных клеток).
- p53 вызывает внутриклеточное увеличение проапоптотических белков семейства Bcl-2 (например, Bax, Bad, Bak).
- Проапоптотические белки увеличивают проницаемость внешней мембраны митохондрий (например, за счет образования мембранного канала гетеродимером Bax / Bad).
- Цитохром c высвобождается из внутренней митохондриальной мембраны и попадает в цитозоль.
- Цитохром c связывается с APAF-1 (фактором активации апоптотической протеазы 1) в цитозоле, образуя колесообразную структуру, известную как апоптосома.
- Комплекс цитохрома с и APAF-1 превращает прокаспазу 9 в активную каспазу 9.
- Каспаза 9 активирует каспазы палача, такие как каспаза 3.
Белки семейства Bcl-2 могут иметь противоположные эффекты: Bad, Bax и Bak обладают проапоптотическим действием, тогда как Bcl-2 и Bcl-xL обладают антиапоптотическим действием.
Аномальная регуляция апоптоза
Гены-супрессоры опухолей, которые регулируют клеточный цикл и гибель клеток, могут мутировать и позволять клеткам оставаться живыми, даже если у них есть аномальные гены, которые могут вызывать рак.
Некроз
Обзор
| Характеристики некроза | ||||
|---|---|---|---|---|
| Тип | Определение | Патофизиология | Внешний вид под микроскопом |
|
|
|
| Разжиженный некроз |
|
| ||
| Фибриноидный некроз 77 77 7 толстые стенки с фрагментами клеточного мусора, сыворотки и фибрина | ||||
| Казеозный некроз | ||||
| Некроз жира |
| |||
| Гангренозный некроз | ||||
Ячеистые включения
Стеатоз
Кальцификация
| Обзор кальцификации | ||
|---|---|---|
| Метастатическая кальцификация | Дистрофическая кальцификация 96 | |
| Описание Диффузная кальцификация нормальной ткани | ||
| Пораженные ткани |
|
|
| Этиология | ||
| Сопутствующие состояния / ткани | 917 тромбосклероз клапана Сердечно-сосудистые заболевания , кальциноз атеросклеротических бляшек
| |
| Содержание кальция в сыворотке | ||
| Гистология | ||
-
Гиалин
- Тип белка, который является эозинофильным при окрашивании H&E (также окрашивается в красный цвет при окрашивании по Ван Гизону) и кажется однородно прозрачным при световой микроскопии.
- Может использоваться для различения внутриклеточного и внеклеточного гиалина.
- Гиалинизация: замена нормальной ткани белками, которые имеют эозинофильный, гомогенный, полупрозрачный вид при окрашивании H&E
Внутриклеточный гиалин
Внеклеточный гиалин
Ссылки
- Фам-Хай Л.А., Хе Х., Фам-Хай К.Свободные радикалы, антиоксиданты при болезнях и здоровье .. Международный биомедицинский журнал: IJBS . 2008; 4 (2): с.89-96.
- Лобо В., Патил А., Фатак А., Чандра Н. Свободные радикалы, антиоксиданты и функциональные продукты питания: влияние на здоровье человека. Обзоры фармакогнозии . 2010; 4 (8): с.118. DOI: 10.4103 / 0973-7847.70902. | Открыть в режиме чтения QxMD
- Игходаро О.М., Акинлое О.А. Антиоксиданты первой линии защиты — супероксиддисмутаза (SOD), каталаза (CAT) и глутатионпероксидаза (GPX): их фундаментальная роль во всей системе антиоксидантной защиты. Александрийский медицинский журнал . 2018; 54 (4): с.287-293. DOI: 10.1016 / j.ajme.2017.09.001. | Открыть в режиме чтения QxMD
- Eltzschig HK, Eckle T. Ишемия и реперфузия — от механизма к трансляции. Нат Мед . 2011; 17 (11): с.1391-1401. DOI: 10,1038 / нм 2507. | Открыть в режиме чтения QxMD
- Kalogeris T, Baines CP, Krenz M, Korthuis RJ. Клеточная биология ишемии / реперфузионного повреждения.. Международный обзор клеточной и молекулярной биологии . 2012; 298 : с.229-317. DOI: 10.1016 / B978-0-12-3-5.00006-7. | Открыть в режиме чтения QxMD
- Ахмад I, Барнетсон Р.Дж., Кришна Н.С. Кератинизирующая плоская метаплазия мочевого пузыря: обзор. Урол Инт . 2008; 81 год (3): с.247-251. DOI: 10,1159 / 000151398. | Открыть в режиме чтения QxMD
- Михалопулос Г.К. Регенерация печени .. J Cell Physiol .2007; 213 (2): с.286-300. DOI: 10.1002 / jcp.21172. | Открыть в режиме чтения QxMD
- Герини Д. Насосы Ca2 + и обменники Na + / Ca2 + .. Биометаллы . 1998; 11 (4): с.319-30.
Все аспекты заживления ран роговицы
Как самая передняя структура глаза, роговица играет важную роль в зрении, механической целостности глаза и иммунологической защите.Он очень структурирован — черта, критически важная для его способности преломлять свет и предотвращать заражение. Однако положение роговицы делает ее уязвимой для различных травм и травм. Правильная функция слезной пленки, век и конъюнктивы имеет важное значение для поддержания роговицы, что делает функцию роговицы уязвимой для нарушения со всех сторон земного шара — и за его пределами. Имеются механизмы заживления роговицы, которые помогают в правильном восстановлении и сохранении структуры роговицы после травмы.А когда они терпят неудачу, доступны терапевтические достижения, направленные на минимизацию долгосрочных осложнений.
Роговица A&P
Типичный эпизод заживления роговичной раны состоит из сложной последовательности событий, в которой задействованы многочисленные типы клеток в соответствующей биохимической среде. Понимание анатомических и физиологических характеристик роговицы — от ее многослойной структуры до ее клеточных соединений — может помочь практикующим врачам лучше понять ключевые клинические моменты этого процесса.
| |
| Рис. 1. Два передних рубца стромы. Щелкните изображение, чтобы увеличить. |
От переднего до заднего, пять знакомых слоев включают роговицу: эпителий, слой Боумена, строму, десцеметовую мембрану и эндотелий. Шестой слой роговицы, так называемый слой Дуа или преддесцеметовый слой, был предложен в качестве необходимого дополнения к списку, но его спорный характер означает, что он пока остается отдельной классификацией. 1
• Эпителий роговицы. Состоящий из пяти-семи слоев клеток и толщиной примерно 50 мкм, эпителий содержит несколько типов клеток, ориентированных слоями от переднего к заднему, параллельно большей структуре роговицы. Конкретные клеточные соединения отвечают за поддержание относительно обезвоженного состояния роговицы, за клеточную связь и обмен материалами, а также за избирательную проницаемость роговицы. Эпителиальные клетки помогают поддерживать стабильную слезную пленку и секретировать базальную эпителиальную мембрану, что имеет решающее значение для заживления роговицы.Клетки эпителия роговицы постоянно переворачиваются, поскольку самые удаленные клетки попадают в слезную пленку. Весь эпителий обновляется примерно за 7-10 дней. Этот процесс ускоряется во время заживления ран и обычно приводит к быстрому заживлению повреждений роговицы, затрагивающих только эпителиальные клетки. 8 (см. «Пристальный взгляд: эпителий роговицы», ).
Подробный обзор: эпителий роговицыПоверхностный слой клеток, составляющих эпителий, имеет толщину примерно в два клеточных слоя и состоит из некератинизированных плоскоклеточных клеток. 2 Эти клетки довольно плоские с многочисленными микроворсинками и микроворсинками, которые увеличивают их общую площадь поверхности, чтобы помочь стабилизировать лежащую сверху слезную пленку. Эпителиальный гликокаликс помогает прикрепить слезы к поверхностным клеткам роговицы, тем самым предотвращая связывание патогенов. 3 Эти поверхностные клетки соединены друг с другом десмосомами, а также плотными контактами, которые в основном состоят из zonula occludens. Плотные соединения помогают обеспечить высокоселективный барьер для веществ в слезной пленке. 4 Межклеточные соединения помогают предотвратить попадание нежелательных веществ в роговицу и помогают поддерживать детургэссенцию роговицы. Второй слой эпителия состоит из двух-трех слоев крыловых клеток, соединенных десмосомами и сращениями. Они прикрепляются спереди к поверхностным клеткам и сзади к базальным клеткам десмосомами, а также соединяются между собой щелевыми соединениями, что позволяет обмениваться молекулами непосредственно между отдельными клетками Базальный клеточный слой состоит из одного слоя столбчатых клеток, которые образуют задний слой. -самый слой эпителия роговицы.Эти клетки прикреплены друг к другу через щелевые контакты, которые играют роль в обеспечении и дифференцировке межклеточной коммуникации, а также через десмосомы. Эти базальные клетки секретируют базальную мембрану, которая делится на переднюю lamina lucida и более заднюю lamina densa. 5 Базальная мембрана эпителия роговицы состоит в основном из коллагенов, ламининов, протеогликанов и нидогенов. 6 Базальная мембрана играет важную роль в клеточных функциях, в том числе участвующих в заживлении, путем контроля связывания факторов роста и их локальных концентраций между слоями клеток.Базальная мембрана базальных клеток прикрепляется через гемидесмосомы к нижележащему слою Боумена, в то время как закрепляющие фибриллы проходят через гемидесмосомы к слою Боумена и вниз в нижележащую строму, где они прикрепляются к закрепляющим бляшкам внеклеточного матрикса, образуя якорный комплекс. 7 |
• Боумановский слой. Этот бесклеточный слой толщиной примерно от 8 до 14 мкм состоит в основном из коллагена и организован в виде беспорядочно расположенных фибрилл диаметром от 20 до 25 нм.Это расположение фибрилл выравнивается по направлению к задней части, поскольку оно смешивается с подлежащей стромой. Слой Bowman обладает высокой устойчивостью к проникновению или повреждению; однако, если он действительно будет поврежден, он не сможет регенерировать, что приведет к его необходимому замещению эпителиальной тканью или тканью стромального рубца.
• Строма. Толщина роговицы во многом обусловлена наличием стромы, которая составляет приблизительно 500 мкм. Этот медиальный слой состоит из организованной сети коллагеновых фибрилл и внеклеточного основного вещества, пористого гидратированного геля, состоящего в основном из агрегатов протеогликана.Кератоциты — специализированные фибробласты — являются основным типом клеток в строме и помогают поддерживать ее целостность. Они также производят коллаген, гликозаминогликаны (ГАГ) и матриксные металлопротеиназы (ММП). Таким образом, точная организация коллагеновых фибрилл в строме роговицы является обязательной для поддержания прозрачности роговицы и соответствующей гидратации стромы.
• Десцеметовая мембрана. Имея толщину всего от 3 до 5 мкм, Descemet’s находится между стромой роговицы и эндотелием, по существу функционируя как базальная мембрана эндотелия роговицы.Десцеметовая мембрана разделена на переднюю пластинку, которая состоит из коллагена, придающего слою эластичные свойства, и заднюю пластинку, которая секретируется эндотелием. Задняя пластинка постоянно продуцируется эндотелием, утолщаясь на протяжении всей жизни. Как и слой Боумена, мембрана Десцемета очень устойчива к травмам, но при повреждении может регенерироваться.
• Эндотелий роговицы. Самый задний слой роговицы состоит из одного слоя многогранных клеток, который поддерживает детургесценцию роговицы с помощью ионных насосов.Базальная поверхность клеток прилегает к десцеметовой мембране, а их апикальная поверхность выстилает переднюю камеру. Хотя эндотелиальные клетки роговицы соединены щелевыми соединениями и плотными соединениями, барьер, образованный этими двумя соединениями, более проницаем, чем передняя поверхность роговицы, что позволяет роговице поглощать питательные вещества из водянистой влаги. Эти клетки не могут делиться или реплицироваться, поэтому, когда они теряются, те, что остаются, используют изменения формы или размера для заполнения пустот в эндотелии.
Заживление эпителиальной раны
Процесс заживления эпителиальной раны роговицы можно разделить на фазы, которые происходят последовательно, но могут перекрываться во времени. Это латентная или лаг-фаза, миграция, пролиферация и повторное прикрепление эпителия.
| |
| Рис. 2. У этого пациента имелся васкуляризированный рубец роговицы. |
• Скрытая фаза. Первая фаза процесса заживления ран роговицы характеризуется клеточным ремоделированием и изменением состава слезы в процессе подготовки к заживлению. 12 Эта фаза приводит к увеличению выработки ферментов (включая ММР-9), которые разрушают поврежденную базальную мембрану эпителия. 4 Матричные металлопротеиназы снижают клеточную адгезию и способствуют увеличению миграции клеток; они также важны в деградации и ремоделировании нормального поддержания внеклеточного матрикса (ЕСМ).Однако чрезмерный уровень этих ферментов может препятствовать заживлению. 13
Латентная фаза включает несколько отдельных этапов и занимает несколько часов: сначала эпителиальные клетки, поврежденные во время травмы, подвергаются апоптозу и попадают в слезную пленку. Затем теряются адгезивные и щелевые соединения в клетках у границы дефекта и нарушаются прикрепления базальных клеток к базальной мембране у краев раны. Эти базальные клетки затем изменяют форму и теряют свои микроворсинки, прежде чем они образуют клеточные расширения, известные как филоподии и ламеллиподии. 9 Затем дефект эпителия покрывается фибронектином, который служит основой для миграции соседних эпителиальных клеток.
• Миграция. Следующая фаза происходит, когда клетки у края раны сглаживаются и распространяются. Филоподии и ламеллиподии отправляются наружу и образуют временные прикрепления к субстрату; затем сократительные элементы тянут клетку вперед к дефекту. 4 Соседние клетки остаются прикрепленными десмосомами и сохраняют свое положение относительно друг друга, скользя по обнаженной области.Вышеупомянутые временные прикрепления затем отщепляются, и филоподии и ламеллиподии снова отправляются вперед, чтобы повторить процесс. Этот цикл продолжается до тех пор, пока дефект не будет полностью закрыт одним слоем ячеек. Обычно процесс занимает от 24 до 36 часов, хотя время может варьироваться в зависимости от местоположения и размера дефекта.
• Распространение. После завершения миграции монослой клеток, покрывающий дефект, разрастается, восстанавливая нормальную толщину эпителия и заполняя дефект.Кратковременные усиливающиеся базальные клетки воспроизводятся посредством митоза, и новые клетки перемещаются внутрь к центру дефекта, а затем вверх, чтобы заполнить его. 9 Клетки превращаются из базальных клеток в клетки крыла, а затем, наконец, клетки плоской поверхности по мере своего движения. Плотные соединения образуются, чтобы восстановить барьерную функцию роговицы, а щелевые соединения, адгезивные соединения и десмосомы образуются между клетками.
• Повторное прикрепление эпителия. Во время заключительной фазы гемидесмосомы снова собираются, чтобы прочно прикрепить эпителиальный слой к субстрату, поскольку закрепляющие фибриллы повторно прикрепляются к закрепляющим бляшкам в строме.Пока базальная мембрана не получила никаких повреждений, этот процесс происходит в считанные дни; Однако если повреждение действительно произошло, окончательное формирование прикреплений и повторное закрепление клеток может занять месяцы или дольше. 8
Заживление стромы
Процесс разрушения стромы ресинтезирует и реорганизует строму роговицы после повреждения стромы и эпителия. Этот процесс, который включает в себя дифференциацию и действия, предпринимаемые кератоцитами, обычно длительный и имеет решающее значение для восстановления прозрачности роговицы.
| |
| Рис. 3. Травматическое истирание роговицы, окрашенное флуоресцеином натрия. Щелкните изображение, чтобы увеличить. |
Первым стромальным событием после повреждения эпителия является апоптоз кератоцитов. На этой стадии растворимые медиаторы из эпителия роговицы вызывают гибель клеток посредством апоптоза лежащих в основе стромальных кератоцитов, в то время как другие кератоциты претерпевают трансформацию в фибробласты и миофибробласты.Кератоциты также увеличивают выработку хемокинов, которые привлекают другие воспалительные клетки в строму роговицы из лимбального кровоснабжения и слезной пленки. Эти воспалительные клетки собирают остатки апоптотических клеток и мусора; кератоциты также могут в конечном итоге стать фибробластами и участвовать в закрытии ран, а также играть роль в регенерации нервов. 13
Считается, что миофибробласты в строме роговицы происходят из кератоцитов под влиянием трансформирующего фактора роста бета (TGF-β) и фактора роста тромбоцитов (PDGF).Миофибробласты создают временный внеклеточный матрикса и создают сократительные силы в попытке закрыть рану; как таковые, они важны для ремоделирования коллагена и внеклеточного матрикса, а также для образования и регрессии помутнения роговицы. 14 Задержка регенерации базальной эпителиальной мембраны (EBM) из-за повреждения, дистрофии или повышенных уровней MMP-2 и MMP-9 может позволить TGF-β и PDGF продолжить попадание в строму роговицы из эпителия, что увековечивает образование миофибробластов.
Продолжающееся присутствие миофибробластов может привести к сильно дезорганизованному ВКМ, что способствует помутнению роговицы и образованию рубцов. Миофибробласты также могут препятствовать надлежащему восстановлению популяции кератоцитов передней стромы, что имеет решающее значение для полного восстановления EBM. 6 Только когда EBM надлежащим образом восстановлен, надлежащие стромальные уровни TGF-β и PDGF устанавливаются, вызывая апоптоз миофибробластов, репопуляцию кератоцитов, устранение аномального ECM и восстановление прозрачности роговицы.
Подавление правильного заживления
Эпителий роговицы поддерживается в сложном балансе, который может быть легко нарушен. Аномалии век или слезной пленки, повреждение нервов роговицы, травмы и инфекции могут нарушить целостность роговицы.
Дефекты эпителия считаются стойкими (PED), если они не поддаются лечению через две недели после начала лечения. PED могут привести к разборке гемидесмосом и деградации слоя Боумена и стромы роговицы.Когда заживление роговицы нарушено, роговица становится особенно уязвимой для повторяющихся дефектов эпителия или повторяющихся эрозий, неоваскуляризации или хронического воспаления стромы и рубцевания (Рисунок 2) .
Рецидивирующие эрозии роговицы (RCE) могут возникать после травмы или при определенных дистрофиях роговицы (Рисунки 3 и 4) . Они изнашивают эпителий роговицы из-за дефектного закрепления эпителиальных клеток, что может происходить из-за неправильного формирования заякоренного комплекса или в результате аномалий EBM.
Нарушения в составе и формировании базальной мембраны, по-видимому, связаны с дистрофией эпителиальной базальной мембраны (EBMD), а также с RCE. Роговица, демонстрирующая EBMD, создает избыточные слои базальной мембраны, которые могут простираться в эпителий роговицы, а не лежать под ним. 15 Эти дополнительные слои базальной мембраны, видимые в виде линий или отпечатков пальцев внутри роговицы, могут препятствовать нормальному перемещению клеток от базального слоя к слоям крыловых и поверхностных клеток эпителия.Кроме того, клетки могут быть захвачены клеточными остатками в базальной мембране с образованием цист. Аномальное наслоение клеток и недостаточная адгезия клеток к подлежащей строме может привести к нарушениям прозрачности роговицы, а также к болезненным RCE. Нарушения в базальной мембране роговицы также могут приводить к увеличению уровней TGF-β1 и PDGF в строме роговицы, что приводит к снижению прозрачности. 16
| |
| Рис.4. Травматическая ссадина роговицы, окрашенная флуоресцеином натрия. |
Неадекватная адгезия эпителия к строме или повреждения, которые повреждают базальную мембрану и нарушают образование гемидесмосом, могут привести к RCE, поскольку плохо прилегающий эпителий легко удаляется путем механического разрушения век. Повышенные уровни MMP-2 и MMP-9 были обнаружены в слезной жидкости пациентов с RCE. 17,18 Увеличение MMP может привести к ненормальной или чрезмерной деградации ECM, препятствуя правильному заживлению ран роговицы и приводя к RCE.Другие состояния и факторы образа жизни, связанные с неправильным заживлением роговицы, включают диабет, нейротрофическое заболевание, заболевание глазной поверхности и курение. Исследование показывает, что изменение сигнальных путей миграции и пролиферации клеток, а также нарушение функции роговичного нерва связаны с замедленным заживлением ран в диабетической роговице. 13
Достижения в лечении
Традиционные цели лечения дефектов эпителия роговицы заключаются в минимизации боли, уменьшении вероятности инфекции и ускорении заживления.Недавний рост понимания патофизиологии RCE и PED привел к новым методам лечения, таким как пероральные тетрациклины, местные стероиды, глазные капли аутологичной сыворотки и наложение или трансплантация амниотической мембраны (AM). Эти процедуры направлены на уменьшение воспаления и оптимизацию среды для заживления.
Документально подтвержденное медицинское использование амниотических оболочек (AM) существует с начала 1900-х годов. Этот материал использовался исследователями в 1940 году в качестве биологической повязки для глазной поверхности, а затем редко упоминался в связи с использованием в офтальмологии до 1995 года, когда исследователи впервые использовали его в качестве хирургического трансплантата для реконструкции глазной поверхности роговицы кролика.В том же году они использовали криоконсервацию для коммерческого изготовления АМ в качестве трансплантата. 11 AM с тех пор пережил возрождение в качестве офтальмологического метода лечения и, как полагают, подавляет воспаление, фиброз и ангиогенез, а также поддерживает заживление ран, помогая миграции и дифференцировке клеток. Исследования также предполагают его использование в качестве противомикробного, противовирусного и обезболивающего средства.
В связи с этим широким спектром действий, влияющих на заживление ран, AM используется для лечения стойких дефектов роговицы и язв, RCE, острых химических или термических ожогов, буллезной кератопатии, частичной недостаточности лимбальных стволовых клеток и для восстановления поверхности ткань конъюнктивы.
Амнион, самый внутренний слой плаценты плода, имеет толщину от 0,02 до 0,5 мм. 20 AM состоит из трех основных слоев: одного слоя кубовидных эпителиальных клеток; толстый слой базальной мембраны; и богатый коллагеном, почти бессосудистый мезенхимальный слой или строма. Строма содержит внутренний компактный слой, средний слой фибробластов и внешний губчатый слой. 21
Эпителиальные клетки AM имеют множество микроворсинок и продуцируют цитокины и другие факторы, участвующие в пролиферации и дифференцировке клеток. 22-25 Они также производят антимикробные пептиды, которые могут снизить риск заражения. 20 Базальная мембрана (BM) AM содержит протеогликаны и другие молекулы, которые поддерживают целостность мембраны и включают различные коллагены, фибронектин, ламинин, фибробласты и факторы роста. 11 BM используется для стимулирования миграции, адгезии и дифференцировки эпителиальных клеток. 26
Строма AM — это богатый коллагеном слой, который обеспечивает прочность на разрыв и содержит большое количество протеогликанов, гликопротеинов и гиалуроновой кислоты (гликозаминогликан).Исследования показывают, что строма AM подавляет передачу сигналов TGF-β, пролиферацию и дифференцировку миофибробластов. 14 Подавление миофибробластов может помочь уменьшить рубцевание, фиброваскулярное врастание и помутнение роговицы. Таким образом, AM может модулировать заживление, способствуя реконструкции тканей, а не образованию рубцов. 20 Строма AM также подавляет воспалительные цитокины и секвестры, инфильтрирующие воспалительные клетки. 25,27
Подробный обзор: лимбальные эпителиальные стволовые клеткиНовые эпителиальные клетки роговицы образуются в базальном слое, единственном митотически активном слое роговицы. 8 Лимбальные эпителиальные стволовые клетки (LESC) расположены рядом с лимбом и обеспечивают поставку базальных клеток, которые позже превращаются в крыловые и поверхностные клетки, мигрируя кпереди. LESC обеспечивают эпителиальные клетки на протяжении всей своей жизни и продуцируют временные амплифицирующие клетки, которые заселяют базальный эпителий периферической роговицы и лимба. 10 Затем клетки мигрируют в центр, пролиферируют и окончательно дифференцируются как центральные эпителиальные клетки. Если LESC повреждены, эпителиальные клетки роговицы могут быть заменены клетками конъюнктивы, что приводит к помутнению роговицы. 11 LESC обычно пролиферируют медленно, но этот процесс ускоряется в ответ на повреждение роговицы, чтобы заселить эпителий роговицы. |
Исследования показывают, что AM может также помочь поддерживать передачу сигналов фактора роста нервов, что, как считается, способствует регенерации нервов. 19,22,25 Кроме того, AM может не экспрессировать большинство основных антигенов гистосовместимости, которые могут привести к отторжению трансплантата. 19,21,27 Более поздние исследования показали, что AM может продуцировать эти антигены только в небольших количествах, что подтверждается отсутствием значительного иммунного ответа, связанного с использованием AM.Это положительное свойство AM может быть связано с иммуносупрессивным эффектом, оказываемым апоптозными AM-клетками, что позволяет использовать AM без необходимости иммуносупрессии. 11,20,21,23,28
Свежий амнион является хорошим источником биологически активных факторов, которые способствуют росту и заживлению ран, пролиферации и миграции эпителиальных клеток, а также ремоделированию внеклеточного матрикса. Ткань можно нагревать или сушить на воздухе, хотя это может привести к потере некоторых биологических свойств ткани.AM также можно лиофилизировать или лиофилизировать, затем стерилизовать гамма-излучением и хранить при комнатной температуре. Было показано, что лиофилизированный AM сохраняет большинство своих физических, биологических и морфологических характеристик. 26 Его также можно хранить на нитроцеллюлозной бумаге и хранить в глицерине для криоконсервации. Криоконсервированный AM может храниться от 12 до 24 месяцев и может сохранять больше первоначальных структурных, физиологических и биохимических свойств мембраны.
Исследования показывают, что эпителиальные клетки AM нежизнеспособны ни в свежих, ни в консервированных препаратах, поскольку вещества выделяются либо из мертвых клеток, либо из поддерживающей базальной мембраны и стромы. 3,23,29 Несмотря на это, хотя некоторые биологические свойства сохраненного AM сохраняются, они, вероятно, сохраняют свою жизнеспособность только в течение ограниченного времени. 24 Криоконсервированные и обезвоженные АМ демонстрируют сопоставимую клиническую эффективность. 26
Трансплантация амниотической мембраны
Клинически AM может использоваться для лечения ран роговицы любого размера и степени тяжести. AM можно использовать в виде трансплантата (в технике вкладок), в виде повязки (в технике накладок) или в комбинации.
Во время процедуры вкладки рана очищается, и AM накладывается эпителием вверх, а затем пришивается на место. AM служит постоянной заменой базальной мембраны; как таковые, соседние эпителиальные клетки реципиента в конечном итоге мигрируют в AM и интегрируют его в роговицу хозяина. 23 AM можно использовать в качестве вкладыша для пациентов с PED или изъязвлениями роговицы или после удаления поражений конъюнктивы.
PED или RCE, которые не реагируют на консервативное лечение искусственными слезами, аутологичной сывороткой и перевязочными контактными линзами, могут особенно выиграть от AM, при этом полное заживление обычно наблюдается в течение одной-двух недель после его применения. 31 Исследование с использованием конфокальной микроскопии для изучения трансплантации амниотической мембраны (AMT) показало, что эпителиальный слой постепенно растворяется в течение двух недель, хотя базальная мембрана и строма трансплантата были обнаружены в течение нескольких месяцев. 29 Фактически, следы трансплантата AM могут оставаться месяцами или годами.
Как часть техники накладок, большая амниотическая мембрана покрывает эпителиальный дефект, а также часть окружающей глазной поверхности подобно бинту контактной линзы.Он защищает нежный эпителий от механического срезания век, действует как барьер для воспалительных клеток и веществ из слезной пленки и способствует гидратации эпителия. Ориентация AM может не иметь особого значения, хотя размещение AM стромы стороной вверх может обеспечить больший контакт с прекорнеальной слезной пленкой. 19 AM пришивается к поверхности глаза, но отслаивается в течение одной-двух недель: можно применить свежий AM, если к этому моменту заживление еще не завершено. 23 AM следует менять еженедельно. Если он остается, он может растворяться в течение недель или месяцев, теряя эффективность.
| |
| Рис. 5. Бесшовный трансплантат амниотической мембраны под бандажной мягкой контактной линзой для лечения нейротрофической язвы. |
AM может использоваться в качестве накладки для состояний, характеризующихся большим количеством воспалений (например, после ожогов или в случае синдрома Стивенса-Джонсона), учитывая его противовоспалительное, антиангиогенное, антиангиогенное действие. — рубцовые и обезболивающие свойства.В этих случаях может быть разумным использовать AM на ранней стадии после начала заболевания, чтобы подавить воспаление и предотвратить рубцовые осложнения.
Техника inlay-onlay используется при обширных изъязвлениях и перфорациях глазной поверхности. Многочисленные более мелкие части AM используются для заполнения дефекта, в то время как более крупная часть используется в качестве накладки. Части накладки защищают детали вкладки и способствуют эпителизации.
Практическое видеоЧтобы узнать больше о клинических аспектах применения амниотической мембраны, вы можете посмотреть это видео с комментариями Натана Лайтзайзера, OD. |
AM может также использоваться в бесшовных технологиях вместе с фибриновым клеем и либо терапевтическими контактными линзами, либо конформером склерального кольца (рис. 5) . 3 Дегидратированный AM доступен в виде безшовного трансплантата, а криоконсервированный AM коммерчески доступен в безшовной форме, закрепленной между двумя несущими кольцами из поликарбоната.
В случаях, когда популяция LESC повреждена или недостаточна, могут возникать PEDs роговицы или поверхность роговицы может заселяться эпителием конъюнктивы, что приводит к хроническому воспалению, рубцеванию, неоваскуляризации или изъязвлению поверхности роговицы.Это делает глаз уязвимым для перфорации или внутриглазной инфекции. 19 Пациентам с полным дефицитом LESC может быть выполнена трансплантация LESC с использованием аутотрансплантата из парного глаза, если он жизнеспособен, или аллотрансплантата от другого донора. 3,22 AM также использовался в качестве биологического субстрата и носителя для культивирования ex vivo лимбальных эпителиальных пластов, предположительно содержащих LESC, что, по мнению исследований, может позволить использовать меньшие трансплантаты LESC для снижения вероятности дефицита LESC для донорский глаз. 19,23 (см. «Более пристальный взгляд: лимбальные эпителиальные стволовые клетки», ).
Совет по кодированиюНе знаете, как кодировать и выставлять счет за использование мембраны? За советом ознакомьтесь с публикацией Coding Connection Джона Румпакиса, OD в этом месяце. |
AM трансплантаты используются для постоянно расширяющегося списка показаний. Их положительные лечебные свойства, низкая иммуногенность и биосовместимость делают их идеальным дополнением к традиционным методам лечения.
Хотя AM анатомически похож на поверхность глаза и его физические характеристики как гибкой биологической мембраны можно непосредственно наблюдать, его клиническая эффективность все еще остается предположительной. 31 Широко распространенные анекдотические сообщения об успешном клиническом применении предполагают, что AM демонстрирует большой потенциал в качестве дополнительного средства к терапевтическому арсеналу, доступному офтальмологам для помощи в заживлении глазной поверхности. Однако необходимы дополнительные исследования, подтверждающие его эффективность в качестве метода первой линии.
Доктор Тара Ли — научный сотрудник Американской академии оптометрии и дополнительный клинический преподаватель Школы оптометрии Калифорнийского университета в Беркли.
Диагностика и лечение атрофического вагинита
ГЛОРИЯ А. БАХМАНН, доктор медицины, и НИКОЛЬ С. НЕВАДУНСКИ, Медицинская школа Роберта Вуда Джонсона, Нью-Брансуик, Нью-Джерси
Am Famician. 2000 15 мая; 61 (10): 3090-3096.
До 40 процентов женщин в постменопаузе имеют симптомы атрофического вагинита.Поскольку это состояние связано с дефицитом эстрогена, оно может возникать у женщин в пременопаузе, которые принимают антиэстрогенные препараты или у которых есть медицинские или хирургические условия, которые приводят к снижению уровня эстрогена. Истонченный эндометрий и повышенный уровень pH влагалища, вызванный дефицитом эстрогена, предрасполагают влагалище и мочевыводящие пути к инфекции и механической слабости. Самые ранние симптомы — уменьшение вагинальной смазки, за которыми следуют другие вагинальные и мочевые симптомы, которые могут усугубляться наложенной инфекцией.После устранения других причин симптомов лечение обычно зависит от замещения эстрогена. Заместительная терапия эстрогенами может проводиться системно или местно, но дозировка и способ доставки должны быть индивидуальными. Влагалищные увлажнители и лубриканты, а также участие в половом акте также могут быть полезны при лечении женщин с атрофическим вагинитом.
Из-за снижения уровня эстрогенов у женщин среднего и старшего возраста часто появляются симптомы атрофического вагинита.По оценкам, от 10 до 40 процентов женщин в постменопаузе имеют симптомы атрофического вагинита, также называемого урогенитальной атрофией.1 Несмотря на распространенность симптомов, только 20-25 процентов женщин с симптомами обращаются за медицинской помощью. для улучшения урогенитального здоровья и качества жизни большого количества пациентов путем выявления и вмешательства в это часто упускаемое из виду и недиагностируемое состояние.
На протяжении жизненного цикла женщины эпителий влагалища претерпевает изменения в ответ на уровень циркулирующего эстрогена.Стимулируемый материнским эстрогеном, эпителий влагалища у новорожденных становится шероховатым и богат гликогеном. В детстве эпителий остается тонким до полового созревания, когда он снова утолщается в результате стимуляции эстрогенами. Стимуляция эстрогеном производит обильное количество гликогена. Лактобациллы Додерлейна зависят от гликогена из отторженных вагинальных клеток3. Молочная кислота, продуцируемая этими бактериями, снижает уровень pH влагалища до 3,5–4,5; это важно для естественной защиты организма от инфекций влагалища и мочевыводящих путей.4 Повышенный уровень pH во влагалище предрасполагает влагалище к инфицированию стрептококками, стафилококками, колиформными бактериями и дифтероидом. 3 После менопаузы уровни циркулирующего эстрогена (в основном эстрадиола) резко снижаются с более чем 120 пг на мл до примерно 18 пг на мл. цитологические преобразования следуют за снижением эстрогена, включая разрастание соединительной ткани, фрагментацию эластина и гиалинизацию коллагена. Эти изменения могут привести к грануляции, трещинам, экхимозам, телеангиэктазам и изъязвлениям.5 Постменопаузальные изменения в составе тканей не ограничиваются половыми путями, но также включают мочевыводящие пути из-за общего эмбриологического происхождения. Эпителий влагалища и уретры зависит от эстрогена и неблагоприятно изменяется в среде, лишенной эстрогена.
Предрасполагающие факторы
Менопауза — основная причина снижения уровня циркулирующего эстрогена; следовательно, это этиология почти всех случаев атрофического вагинита. У женщин без менопаузы выработка эстрогена яичниками может быть прервана лучевой терапией, химиотерапией, иммунологическими нарушениями и овариэктомией.Послеродовое снижение уровня эстрогена сопровождается потерей эстрогена в плаценте и антагонистическим действием пролактина на выработку эстрогена во время лактации. Побочные эффекты антиэстрогенных препаратов, включая медроксипрогестерон (Провера), тамоксифен (Нолвадекс), даназол (Данокрин), лейпролид (Лупрон) и нафарелин (Синарел), также являются причинами атрофического вагинита.6 Происходит усиление тяжести симптомов. у женщин с естественным пременопаузальным дефицитом эстрогена, курящих сигареты, не рожавших естественным путем или у которых уровень эстрогена не колеблется.3,7,8 Более легкая атрофия встречается у женщин в постменопаузе, которые участвуют в половом акте, имеют более высокий уровень андрогенов и не подвергались вагинальной хирургии (таблица 1). 3,6–9
Посмотреть / распечатать таблицу
ТАБЛИЦА 1Факторы, которые Повышает риск развития атрофического вагинита
| Менопауза | ||
| Снижение функции яичников | ||
| Лучевая терапия | | |
Овариэктомия
Послеродовая потеря эстрогена из плаценты
Повышенный уровень пролактина во время кормления грудью
Даназол (Данокрин)
Медроксипрогестерон (Провера)
Лепролид (Сина1694
Лупролид (Лупарелин
79
Естественный дефицит эстрогенов до наступления менопаузы3
Курение сигарет7
Вагинальное недородство
Не меняющиеся уровни эстрогена
| Менопауза | |||||||
| Снижение функции яичников | |||||||
| Лучевая терапия | терапия | ||||||
| Иммунное расстройство | |||||||
| Офорэктомия | |||||||
| Послеродовая потеря эстрогена плаценты | |||||||
|
| 00 | 00 | 00 | 00 | 00 | 00 9174 9174 | |
|---|---|---|---|---|---|---|---|
| Тамоксифен (Нолвадекс) | |||||||
| Даназол (Данокрин) | |||||||
| Медроксипрогестерон | 94 194 | Нафарелин (Синарел) | |||||
| Естественный дефицит эстрогенов до наступления менопаузы3 | |||||||
| Курение сигарет7 | |||||||
| 697 | 9 0004 Неизменяющиеся уровни эстрогена8|||||||
| Прекращение половой жизни9 | |||||||
Признаки и симптомы
Как правило, до появления симптомов атрофического вагинита требуется долгосрочное снижение стимуляции эстрогеном.Уменьшение вагинальной смазки является ранним признаком гормональной недостаточности.10 Симптомы со стороны половых органов включают сухость, жжение, диспареунию, потерю влагалищных выделений, лейкорею, зуд вульвы, чувство давления, зуд и желтые выделения с неприятным запахом3,6,11. дискомфорта уретры, частота, гематурия, инфекция мочевыводящих путей, дизурия и стрессовое недержание мочи могут быть более поздними симптомами вагинальной атрофии (таблица 2) .3,6,10,11 Все симптомы атрофического вагинита могут усугубляться одновременным заражением кандидозом, трихомониазом или бактериальный вагиноз.Со временем отсутствие вагинальной смазки часто приводит к сексуальной дисфункции и связанным с ней эмоциональным расстройствам.
Просмотр / печать таблицы
ТАБЛИЦА 2Симптомы атрофического вагинита
| Генитальный | ||||
| Сухость | ||||
| 00 9 00 9 00 00 00 00 00 | 9 | |||
| Диспареуния | ||||
| Жгучая лейкорея | ||||
| Вульвар почесать 9174 927 927 927 927 927 927 927 927 927 927 927 927 9174 927 выделения | ||||
| Мочевыводящие | ||||
| Дизурия | ||||
| Гематурия | ||||
| Стрессовое недержание мочи | ||||
Симптомы атрофического вагинита
|
| ||
| 9165 | 00 7 | |
|---|---|---|
| Жжение | ||
| Диспареуния | ||
| Горящий бели5999169 | 00 |
|---|
1 927 927 927 927 927
Желтые выделения с неприятным запахом
Мочевые
Дизурия
Частота мочеиспускания
Инфекция мочевыводящих путей
Стрессовое недержание мочи
Диагноз
EX2000 не важен для диагноза (или только диагноз атрофического вагинита) у пациентки в постменопаузе с жалобами на урогенитальные органы.В анамнезе пациента следует обратить внимание на экзогенные агенты, которые могут вызывать или еще больше усугублять симптомы. Духи, порошки, мыло, дезодоранты, прокладки для трусов, спермициды и лубриканты часто содержат раздражающие соединения.6 Кроме того, плотно прилегающая одежда и длительное использование промежностных прокладок или синтетических материалов могут ухудшить симптомы атрофии12 (Таблица 3) .6,12Просмотреть / распечатать таблицу
ТАБЛИЦА 3Дифференциальная диагностика атрофического вагинита
| Кандидоз | ||
| Бактериальный вагиноз | ||
| Парфюмерия | ||
| Порошки | ||
| Дезодоранты | 00 | 00 91679271719277 | | | Мыло | | Спермициды | | Смазочные материалы | | Плотно прилегающая или синтетическая одежда | Бактериальный вагиноз | Трихомониаз | Контактное раздражение или реакция на: | 9 9 9 9 9 9 9 | | Дезодоранты | | Прокладки для трусиков | | Промежуточные прокладки | 74000 00 927mic89 | | | Смазочные материалы | | Облегающая или синтетическая одежда |
|---|---|---|
При осмотре будут очевидны несколько признаков атрофии влагалища.Атрофический эпителий выглядит бледным, гладким и блестящим. Часто может присутствовать воспаление с пятнистой эритемой, петехиями и повышенной рыхлостью. Наружные гениталии следует исследовать на предмет снижения эластичности, тургора кожи, разреженности лобковых волос, сухости половых губ, дерматозов вульвы, поражений вульвы и сращения малых половых губ3,6 (Рисунок 1). Будет очевиден интроитальный стеноз шириной менее двух пальцев и уменьшенная глубина влагалища; если эти состояния не диагностируются до введения расширителя, тазовое обследование вызовет значительную боль.Рыхлый и плохо шероховатый эпителий влагалища более подвержен травматическим повреждениям. Экхимозы и небольшие разрывы в периинтроитальном и заднем отделах фуршета также могут повторяться после коитуса или во время осмотра зеркала. Вагинальный осмотр или половая жизнь могут привести к вагинальному кровотечению или кровянистым выделениям. Признаки раздражения вульвы, вызванные недержанием мочи, также могут быть выявлены при гинекологическом осмотре. Цистоцеле, полипы уретры, карункул уретры, выворот слизистой оболочки уретры, пролапс тазовых органов и ректоцеле часто сопровождают атрофический вагинит3 (Таблица 4).3,6
Просмотр / печать Рисунок
РИСУНОК 1.
Наружные гениталии 67-летней женщины, у которой естественная менопауза в течение двух лет и которая не проходит заместительную терапию эстрогенами. Обратите внимание на потерю полноты губ и вульвы, бледность эпителия уретры и влагалища и уменьшение влажности влагалища.
РИСУНОК 1.
Наружные гениталии 67-летней женщины, у которой естественный менопаузальный период в течение двух лет, и которая не получает заместительной терапии эстрогенами.Обратите внимание на потерю полноты губ и вульвы, бледность эпителия уретры и влагалища и уменьшение влажности влагалища.
Просмотр / печать таблицы
ТАБЛИЦА 4Физические признаки атрофического вагинита
| Генитальный | ||
| Бледный, гладкий или блестящий эпителий влагалища 17411 | ||
| Редкость лобковых волос | ||
| Сухость половых губ | ||
|
| 94 927 927 927 + | рыхлый, unrugated эпителий | + 91 694 тазовых органов Выпадение | + 91 694 ректоцеле | + 91 694 вульвы дерматозы | + 91 694 вульвы lesio нс |
| вульвы патч эритема | ||
| Петехии эпителия | ||
| уретральный | ||
| уретральный сосочек | ||
| Выворот Слизистая оболочка уретры | ||
| Цистоцеле | ||
| Полипы уретры | ||
| 94 911 927 911 927 927 927 911 911 927 911 927 927 927 | ||
Физические признаки атрофического вагинита
| Генитальный | |||
| Бледный, гладкий или блестящий эпителий влагалища 917urg11 | |||
| 1 | из | кожи 711 | |
| Редкость лобковых волос | |||
| Сухость половых губ | |||
| Сращение малых половых губ tenosis | |||
| рыхлый, unrugated эпителий | |||
| тазовых органов Выпадение | |||
| ректоцеле | |||
| вульвы дерматозы | |||
| вульвы поражений | |||
| Эритема пятна вульвы | |||
| Петехии эпителия | |||
| Уретральная | |||
| 1 1 | |||
| 1 1 927 927 927 9279 разрастание слизистой уретры | |||
| Цистоцеле | |||
| Полипы уретры | |||
| 00 | 00 91719114 | ||
|---|---|---|---|
ЛАБОРАТОРНЫЕ РЕЗУЛЬТАТЫ
Лабораторные диагностические исследования, включая уровни гормонов сыворотки и мазок Папаниколау, могут подтвердить наличие урогенитальной атрофии (Рисунки 2 и 3; Таблица 5).3,13 Цитологическое исследование мазков из верхней трети влагалища показывает увеличение доли парабазальных клеток и снижение процента поверхностных клеток. Повышенный уровень pH (уровень pH в постменопаузе превышает 5), который контролируется полоской pH в своде влагалища, также может быть признаком атрофии влагалища. Кроме того, вагинальная ультрасонограмма слизистой оболочки матки, которая демонстрирует тонкий эндометрий размером от 4 до 5 мм, означает потерю адекватной эстрогенной стимуляции.13 При микроскопической оценке потеря поверхностных клеток очевидна при атрофии, но также могут быть признаки инфекции. при трихомонаде, кандидозе или бактериальном вагините.
Просмотр / печать Рисунок
РИСУНОК 2.
Нормальный мазок Папаниколау с шейки матки, демонстрирующий плоскоклеточные клетки из поверхностного и промежуточного слоев эпителия. Клетки имеют обильную цитоплазму и низкое ядерно-цитоплазматическое соотношение. По мере того, как клетки созревают по направлению к поверхности, цитоплазма становится ороговевшей, приобретая розовый цвет, а ядро становится маленьким и конденсированным, отражая пикноз.
РИСУНОК 2.
Нормальный мазок Папаниколау из шейки матки, демонстрирующий плоскоклеточные клетки из поверхностного и промежуточного слоев эпителия. Клетки имеют обильную цитоплазму и низкое ядерно-цитоплазматическое соотношение. По мере того, как клетки созревают по направлению к поверхности, цитоплазма становится ороговевшей, приобретая розовый цвет, а ядро становится маленьким и конденсированным, отражая пикноз.
Просмотр / печать Рисунок
РИСУНОК 3.
Мазок Папаниколау демонстрирует атрофический вагинит с незрелыми (парабазальными) плоскоклеточными эпителиальными клетками с увеличенными ядрами на фоне базофильных гранулярных обломков и воспалительного экссудата.Присутствуют характерные аморфные базофильные структуры округлой формы («синие пятна»).
РИСУНОК 3.
Мазок Папаниколау демонстрирует атрофический вагинит с незрелыми (парабазальными) плоскоклеточными эпителиальными клетками с увеличенными ядрами на фоне базофильных гранулярных обломков и воспалительного экссудата. Присутствуют характерные аморфные базофильные структуры округлой формы («синие пятна»).
Просмотреть / распечатать таблицу
ТАБЛИЦА 5Лабораторная диагностика атрофического вагинита
| Лабораторный тест | Положительный результат |
|---|---|
| Мокрый препарат / цитологический мазок клеток из верхней части 5 5 влагалища Атрофические цитологические изменения, включая увеличение доли парабазальных клеток | |
| Ультрасонография слизистой оболочки матки | Выстилка матки, демонстрирующая тонкость эндометрия между 4 и 5 мм 13 | 9
| Вагинальный pH | Повышение pH выше нормального уровня в постменопаузе (pH выше 5) 3 |
| Микроскопия | Устранение диагноза |
Лабораторная диагностика атрофического вагинита
| Лабораторный тест | Положительный результат | |
|---|---|---|
| Мокрый препарат / цитологический мазок клеток из верхней трети влагалища | Атрофические изменения в пропорции парабазальные клетки | |
| Ультрасонография слизистой оболочки матки | Внутренняя слизистая оболочка матки демонстрирует тонкость эндометрия от 4 до 5 мм 13 | |
| Уровень гормона в сыворотке крови | 0 Низкая концентрация гормона в сыворотке | 0 Строген ≤ 4.5 |
| Вагинальный pH | Повышение pH выше нормального уровня в постменопаузе (pH более 5) 3 | |
| Микроскопия | Устранение диагноза |
Лечение
ЗАМЕНА ЭСТРОГЕНОВ
Поскольку отсутствие циркулирующих естественных эстрогенов является основной причиной атрофического вагинита, заместительная гормональная терапия является наиболее логичным выбором лечения и доказала свою эффективность в восстановлении анатомии и устранении симптомов .Замещение эстрогена восстанавливает нормальный уровень pH, утолщает и реваскуляризует эпителий. Адекватная заместительная терапия эстрогенами увеличивает количество поверхностных клеток.3 Терапия эстрогенами может облегчить существующие симптомы или даже предотвратить развитие урогенитальных симптомов, если ее начать во время менопаузы. Противопоказания к терапии эстрогенами включают эстроген-чувствительные опухоли, конечную стадию печеночной недостаточности и тромбоэмболизацию, связанную с эстрогеном, в анамнезе. Побочные эффекты эстрогеновой терапии включают болезненность груди, вагинальное кровотечение и небольшое повышение риска эстроген-зависимого новообразования.14 Повышенный риск развития карциномы и гиперплазии эндометрия окончательно связан с беспрепятственным приемом экзогенного эстрогена.15 Факторы, определяющие степень повышенного риска, включают продолжительность, дозировку и способ доставки эстрогена. Пути введения включают пероральный, трансдермальный и интравагинальный. Частота приема может быть непрерывной, циклической или симптоматической. Количество эстрогена и продолжительность времени, необходимого для устранения симптомов, сильно зависят от степени атрофии влагалища и различаются у разных пациентов.
Было показано, что системное введение эстрогенов оказывает терапевтический эффект на симптомы атрофического вагинита. Дополнительные преимущества системного введения включают уменьшение потери костной массы в постменопаузе и облегчение вазомоторной дисфункции (приливы). Стандартные дозы системного эстрогена, однако, не могут устранить симптомы атрофического вагинита у 10–25 процентов пациентов16. Для облегчения симптомов может потребоваться системный эстроген в более высоких дозах. Некоторым женщинам требуется одновременное введение вагинального препарата эстрогена, применяемого местно.Для полного устранения сухости может потребоваться до 24 месяцев терапии; однако некоторые пациенты не полностью реагируют даже на этот режим лечения.10
Другие варианты лечения включают трансвагинальную доставку эстрогена в виде кремов, пессариев или гормонального кольца (Estring). Лечение низкими дозами трансвагинального эстрогена оказалось эффективным для облегчения симптомов, не вызывая значительной пролиферации вагинального эпителия.2,12,14,17 Уровень pH мочеполовой системы также снижается, что приводит к снижению частоты инфекций мочевыводящих путей.Скорость абсорбции увеличивается с увеличением продолжительности лечения из-за усиления кровоснабжения обработанного эпителия. Преимущество трансвагинального лечения может заключаться в снижении риска карциномы эндометрия, поскольку для устранения урогенитальных симптомов требуется меньшее количество гормона. Негативные эффекты трансвагинального лечения включают неприязнь пациента к вагинальным манипуляциям, меньшую профилактику потери костной массы в постменопаузе и вазомоторную дисфункцию, снижение контроля абсорбции вагинальных кремов по сравнению с пероральными и трансдермальными введениями, а также нерегулярные интервалы лечения, из-за которых пациенты могут забыть о назначении лечения. .6
Трансвагинальные кольца предлагают удобство, постоянство концентрации гормонов в кровотоке и терапевтическую ценность, эквивалентную кремам, без необходимости частого применения. Управление дозировкой гормона осуществляется путем изменения площади поверхности кольца. Симптомы атрофического вагинита облегчаются (с дозировкой от 5 до 10 мкг в 24 часа) без стимуляции пролиферации эндометрия, тем самым устраняя необходимость добавления в схему лечения прогестагена, противоположного этому действию.18 Большинство пациентов могут без труда удалить и снова вставить кольца. можно носить во время коитуса.
УВЛАЖНЯЮЩИЕ И СМАЗОЧНЫЕ СРЕДСТВА
Увлажняющие и смазывающие вещества могут использоваться в сочетании с заместительной терапией эстрогенами или в качестве альтернативного лечения.17 Некоторые пациенты предпочитают не принимать заместительные гормоны, или у них могут быть медицинские противопоказания или гормональные побочные эффекты. Пациентам, которые не хотят использовать эстроген, не следует использовать увлажняющие средства, содержащие женьшень, поскольку они могут обладать эстрогенными свойствами19. Увлажняющие средства помогают поддерживать естественную секрецию и комфорт при половом акте.Продолжительность действия обычно составляет менее 24 часов.
Сексуальная активность
Сексуальная активность — это полезный рецепт для женщин в постменопаузе, у которых имеется существенно эстрогенизированный вагинальный эпителий. Было показано, что он способствует эластичности и податливости влагалища, а также стимулирует лубрикантную реакцию на сексуальную стимуляцию. Женщины, которые участвуют в сексуальной активности, сообщают о меньшем количестве симптомов атрофического вагинита и при вагинальном обследовании имеют меньше признаков стеноза и сжатия по сравнению с сексуально неактивными женщинами.Отрицательная взаимосвязь существует между половым актом, включая мастурбацию, и симптомами атрофии влагалища.9
Поскольку не было показано, что существует положительная взаимосвязь между уровнем эстрогена и сексуальной активностью, не предполагается, что коитус восстанавливает или поддерживает эстроген у женщин в постменопаузе. Наличие положительной взаимосвязи между половым актом и гонадотропинами и андрогенами указывает на важность этих соединений для здорового вагинального эпителия при снижении уровня эстрогена.9 Все сексуально активные женщины должны принимать соответствующие меры предосторожности против заболеваний, передающихся половым путем, включая вирус иммунодефицита человека.
Заключительный комментарий
Атрофия влагалища не обязательно является неизбежным следствием менопаузы или других событий, которые приводят к длительной потере эстрогена. Активная диагностика и вмешательство могут предотвратить развитие атрофического вагинита или устранить существующие симптомы. Осведомленность о множестве вариантов замены эстрогена, а также альтернативных методов лечения значительно увеличивает способность врача назначать лечение, соответствующее физическим потребностям и образу жизни пациента.При определенных обстоятельствах поощрение сексуальной активности также является важным источником нефармакологического лечения, о котором многие пациенты могут не быть проинформированы. По иронии судьбы, продолжающиеся половые отношения могут повысить способность женщины вести здоровую сексуальную жизнь после менопаузы, поддерживая физиологическую среду, защищающую от атрофических изменений.
Механизмы поражения эпителия дыхательных путей: эпителиально-мезенхимальные взаимодействия в патогенезе астмы
Цели этой статьи — понять недостатки в представлении об астме исключительно как об атопическом заболевании, получить представление об эпителиальных аномалиях астмы и о том, как они взаимодействуют с окружающей средой, а также рассмотреть астму как заболевание, связанное с перестройкой стенки дыхательных путей, которое задействует активацию эпителиальной мезенхимальной трофической единицы (EMTU).Кроме того, на основе этих разработок определены возможные новые терапевтические цели для лечения хронической астмы.
Механизмы поражения эпителия дыхательных путей
Давно признано, что воспаление дыхательных путей типа 2 из Т-хелперов лежит в основе дисфункции дыхательных путей при астме. Атопия также является ключевым признаком этого заболевания, но составляет <40% от общего риска у населения. Основываясь на тщательной патологической и функциональной оценке хронической астмы, авторы предполагают, что измененная эпителиально-мезенхимальная связь также имеет фундаментальное значение для патогенеза заболевания.Ряд факторов способствует утолщению гиперчувствительных дыхательных путей, что обеспечивает идеальную микросреду для стойкости Th3-опосредованного воспаления, включая более восприимчивый эпителий к травмам, замедленное восстановление эпителия, измененную траекторию восстановления эпителия до фенотипа, выделяющего слизь, образование факторов роста, которые управляют пролиферацией и дифференцировкой мезенхимальных клеток в направлении увеличения отложения матрикса и гладких мышц, а также продукции нервных и сосудистых факторов роста.Цитокины и медиаторы, полученные из инфильтрирующих воспалительных клеток, взаимодействуют с этим EMTU, чтобы усилить и продлить ответы. «Ремоделирующие» изменения астмы наблюдались при детской астме и действительно могут предшествовать развитию болезни. Недавнее признание того, что атопия как таковая не является ключевым фактором в инициировании астмы (хотя она важна для обострения болезни), предполагает, что взаимодействия генов и окружающей среды, включая процессы, аналогичные тем, которые происходят при морфогенезе ветвления, имеют решающее значение для полной астмы. фенотип развиваться.
Введение в астму
Недавний семинар национальных институтов здравоохранения и рабочая группа Европейского респираторного общества пришли к выводу, что необходима дополнительная работа, чтобы понять механизмы тяжелой и хронической астмы 1, 2. В отдельности Th3-опосредованное воспаление недостаточно, чтобы вызвать астму, а скорее астматическое. Состояние возникает в результате сочетания повышенной восприимчивости эпителия бронхов к повреждению и длительного восстановления тканей, включая дисбаланс в ответах на лиганды рецептора тирозинкиназы, такие как фактор роста эпителия (EGF) и трансформирующий фактор роста (TGF) -β и Th3 цитокины.Они изменяют межклеточную связь между эпителием и подлежащей оболочкой фибробластов, вызывая активацию EMTU, и таким образом распространяют и усиливают структурные изменения в дыхательных путях, создавая микросреду для поддержания хронического воспаления.
Астма: динамическое расстройство воспаления и ремоделирования
Астма — это воспалительное заболевание дыхательных путей с участием Т-клеток, тучных клеток и эозинофилов, характерное для Th3-подобного ответа.Однако одно лишь воспаление не объясняет многих особенностей этого хронического и рецидивирующего заболевания. Хотя атопия является важным фактором риска развития астмы, в общей популяции на нее приходится <40% приписываемого риска заболевания этим заболеванием 3. Предполагается, что эозинофилы играют центральную роль в патогенезе заболевания, однако выводы, что антибиотик -интерлейкин (ИЛ) -5-блокирующее моноклональное антитело 4 и рекомбинантный человеческий ИЛ-12 5 не оказывают значительного влияния на аллерген-индуцированные ранние или поздние фазовые ответы, несмотря на заметное снижение количества эозинофилов в кровотоке и дыхательных путях, что поставило вопрос о первичности этой клетки при астме.Таким образом, будучи ассоциированным с астмой, атопия и эозинофилия дыхательных путей, по-видимому, не являются критическими требованиями для проявления болезни. Генетические исследования продемонстрировали, что атопия и гиперреактивность бронхов (BHR) имеют разные модели наследования 6. Эти результаты предполагают, что местные операционные факторы играют важную роль в предрасположенности людей к астме, и дают объяснение эпидемиологическим данным, определяющим воздействие загрязняющих веществ 7, диета 8 и респираторная вирусная инфекция 9, которые все усиливают окислительный стресс в дыхательных путях.
Морфометрия показала, что утолщение астматических дыхательных путей является причиной значительного компонента BHR и чрезмерного сужения дыхательных путей, наблюдаемого при установленной астме. При умеренном / тяжелом заболевании структурные изменения и ЧСС плохо поддаются кортикостероидам 10. Этот недостаток кортикостероидов отражен в результатах недавнего исследования Европейской сети по изучению тяжелой астмы, которое показало, что эти пациенты демонстрируют значительно сниженные качества. of life 11, компонент фиксированной обструкции дыхательных путей и имеет четкие доказательства ремоделирования стенки дыхательных путей 12.У таких пациентов были обнаружены признаки стойкого обновления матрикса. Ремоделирование дыхательных путей при астме у взрослых также объясняет ускоренное снижение функции легких, наблюдаемое с течением времени. 13. Исследование Программы управления детской астмой (CAMP) у детей в возрасте 5–11 лет показало, что первоначальный положительный эффект ингаляционных кортикостероидов на Постбронходилатирующее улучшение функции дыхательных путей, наблюдавшееся в течение первого года лечения, было потеряно в течение следующих 3 лет. 14. Более недавнее исследование «Лечение ингаляционными стероидами как регулярная терапия в раннем исследовании астмы» (START) проводилось на очень большой популяции впервые диагностированных астматиков. ясно продемонстрировал эффективность ингаляционного будесонида по многим критериям исхода астмы, но опять же, очень ограниченное или нулевое влияние на естественное течение, измеренное с помощью объема форсированного выдоха после бронходилататора за одну секунду в течение 3 лет 15.Лучше всего это объясняется перестройкой дыхательных путей, нечувствительной к кортикостероидам. Недавнее исследование биопсии выявило ремоделирование ткани как ранний и постоянный компонент детской астмы с пролиферацией фибробластов и отложением коллагена в субэпителиальной ретикулярной пластинке, что имеет большее диагностическое значение, чем тканевая эозинофилия 16. Хотя ремоделирование считалось вторичным по отношению к длительной болезни. стоячое воспаление, исследования биопсии у маленьких детей показали реструктуризацию тканей за 4 года до появления симптомов 17, что указывает на процессы, которые начинаются на ранней стадии развития астмы и происходят параллельно или могут быть обязательными для установления стойкого воспаления .
Восприимчивость эпителия к повреждению и фенотип восстановления при астме
Нормальный бронхиальный эпителий представляет собой многослойную структуру, состоящую из столбчатого слоя, поддерживаемого базальными клетками, которые служат физическим и химическим барьером для внешней среды. При астме эпителий демонстрирует множество признаков повреждения и активации, а также метаплазии бокаловидных клеток. Эпителиальный стресс проявляется в широко распространенной активации ядерного фактора транскрипции-κB 18, 19, белков-активаторов ( e.грамм. активаторный белок-1) 20 и преобразователь сигнала и активация транскрипции (STAT) -111, а также за счет повышенной экспрессии белков теплового шока 22 и ингибитора циклин-зависимой киназы, p21 waf 23. Измененный эпителий также становится важный источник аутакоидных медиаторов, хемокинов и факторов роста 24, поддерживающих продолжающееся воспаление.
Чувствительность эпителия
Хотя сообщается, что разрушение эпителия бронхиального эпителия при астме увеличивается, было высказано предположение, что это является артефактом 25.Однако находки повышенной экспрессии рецептора эпидермального фактора роста (EGFR) 26 и эпителиальной изоформы CD44 27 показывают, что повреждение произошло in vivo . Экспрессия EGFR и CD44 при астме увеличивается пропорционально тяжести заболевания, и это очевидно по всему эпителию, что позволяет предположить, что повреждение широко распространено 26, 27. Сверхэкспрессия EGFR также нечувствительна к действию кортикостероидов и положительно коррелирует с толщиной ретикулярной пластинки. 26, что обеспечивает основу для связи повреждения эпителия с лежащей в основе реакцией ремоделирования.Степень отслаивания эпителия при астме не наблюдается при других воспалительных заболеваниях, таких как хроническая обструктивная болезнь легких (ХОБЛ). При ХОБЛ эпителий становится многослойным и претерпевает плоскоклеточную метаплазию, в то время как нижележащая сетчатая пластинка остается нормальной. Хотя возможно, что эти различия отражают тип присутствующего воспаления, эозинофилия дыхательных путей наблюдается в отсутствие астмы 28, и нейтрофилы могут доминировать в воспалении при тяжелой астме, как при ХОБЛ 12.
В биопсии бронхов заметно усиленное иммуноокрашивание астматических столбчатых (но особенно не базальных) эпителиальных клеток для p85, продукта расщепления каспазы-3 полимеразы поли (аденозиндифосфат-рибозы), что указывает на усиление апоптоза эпителия при этом заболевании 29 .Хотя эти гистологические исследования выявляют эпителиальные различия между астматиками и здоровыми людьми, они не могут различить, являются ли изменения причиной или следствием воспаления. Чтобы решить эту проблему, авторы создали первичные культуры с использованием эпителиальных клеток бронхов, взятых из дыхательных путей нормальных и астматических субъектов, чтобы сравнить ответы в идентичных условиях in vitro . Хотя в оптимальных условиях роста не было обнаружено никакой разницы в скорости пролиферации этих культур, в состоянии покоя из-за истощения фактора роста эти клетки из дыхательных путей астмы проявляют значительно большую чувствительность к индуцированному оксидантами апоптозу при нормальном апоптотическом ответе. к дезоксирибонуклеиновой кислоте (ДНК) и ингибитору синтеза рибонуклеиновой кислоты, актиномицину D 29.Эта восприимчивость к окислителям вряд ли является вторичным эффектом воспаления дыхательных путей, поскольку она сохраняется в течение нескольких поколений in vitro . Поскольку эпидемиологические исследования 30, 31 выявили множественные взаимодействующие факторы риска астмы, включая ингаляционные загрязнители и диеты с низким содержанием антиоксидантов, авторы настоящей статьи предполагают, что влияние оксидантов окружающей среды на чувствительный эпителий обеспечивает вероятный пусковой механизм для индукции активации эпителия и повреждение при астме.После инициирования возникающий в результате приток воспалительных клеток вызывает вторичное повреждение за счет производства эндогенных активных форм кислорода, что приводит к хроническому повреждению тканей и стойкому воспалению.
Недавно было показано, что эпителиальная экспрессия нейтрофильных хемоаттрактантов IL-8 и воспалительного белка макрофагов (MIP) -1α увеличивается при тяжелой астме, и что их появление коррелирует с повышенной экспрессией EGFR как маркера повреждения эпителия. Дальнейшие исследования in vitro и показали, что обработка первичных бронхиальных эпителиальных клеток окислителем или EGF усиливает высвобождение MIP-1α или IL-8 соответственно 32.Таким образом, помимо легкого перемежающегося заболевания, которое тесно связано с атопией, восприимчивость эпителиального барьера к действию различных компонентов вдыхаемой среды играет ключевую роль в определении астматического фенотипа.
Остановка роста и восстановление эпителия
Исследования in vitro указывают на центральную роль активации EGFR в восстановлении бронхиального эпителия после повреждения, поскольку EGF является митогеном для эпителиальных клеток бронхов 23, механическое повреждение вызывает быстрое фосфорилирование EGFR независимо от наличия экзогенный лиганд 26, и закрытие раны усиливается за счет EGF, но не за счет неродственного лиганда, фактора роста кератиноцитов 26.Хотя EGF является мощным митогеном, увеличение эпителиального EGFR при астме не сопровождается повышенной пролиферацией для замещения столбчатых клеток, которые были потеряны в результате расщепления каспазой молекул межклеточной адгезии 23, 33. Это контрастирует с гиперпролиферативным состоянием эпителий, наблюдаемый при ХОБЛ. Хотя исследования с первичными культурами нормальных и астматических эпителиальных клеток бронхов показали схожую скорость пролиферации при поддержании в среде с добавлением экзогенного EGF 29, авторы обнаружили потенциальный механизм снижения EGFR-опосредованной пролиферации in vivo в отношении циклин-зависимой Ингибитор киназы, p21 waf , который сверхэкспрессируется как в базальных, так и в столбчатых эпителиальных клетках в биоптатах бронхов пациентов с тяжелой астмой 23.В то время как при легкой форме астмы p21 waf в цитоплазматическом месте защищает столбчатые клетки от окислительного повреждения, при более тяжелом заболевании он экспрессируется в базальных клетках в ядерной локализации, где он служит контрольной точкой при переходе из G1 в S-фазу. клеточный цикл, вызывающий остановку роста. Это позволяет завершить репарацию ДНК до перехода в S-фазу или, если повреждение необратимо, напрямую выйти из клеточного цикла и активировать апоптоз. Таким образом, решение выжить или вступить в апоптоз связано.Лиганды EGFR являются детерминантами судьбы эпителиальных клеток благодаря их способности действовать как факторы выживания 34, которые защищают от проапоптотических стимулов, и как митогены, которые сигнализируют о прогрессировании клеточного цикла. Хотя способность EGF защищать от индуцированного окислителем апоптоза in vitro еще не охарактеризована, исследования на животных моделях показали, что EGF может защищать от вызванного дымом повреждения трахеи у овец 35.
Поскольку экспрессия p21 waf сильно индуцируется оксидантным стрессом 25, открытие, что астматические эпителиальные клетки бронхов более восприимчивы к окислительному повреждению, дает одно из объяснений его высокой экспрессии в астматическом эпителии.Однако p2I waf также индуцируется антипролиферативными факторами роста TGF-β 1 и TGF-β 2 , уровни которых повышены при астме, ХОБЛ 36 и в ответ на повреждение эпителия in vitro 26, 37 Поскольку активация митоген-активируемой протеинкиназы (MAPK) митогенами, такими как EGF, антагонизирует передачу сигналов TGF-β 38, общая судьба эпителия будет отражать интеграцию сигналов выживания и пролиферации, обеспечиваемых EGF-подобными факторами роста, и уравновешивающим эффектом проапоптоза. и антипролиферативные сигналы, вызванные повреждением и членами семейства TGF-β, включая костные морфогенные белки.Хотя экспрессия лигандов EGFR 39 в астматическом бронхиальном эпителии не изменяется по сравнению с нормальным контролем, при ХОБЛ наблюдается заметно повышенная продукция TGF-α. Таким образом, при ХОБЛ повышенная активация EGFR с помощью EGF и аналогичных лигандов защищает от повреждений, вызванных сигаретным дымом, и отменяет антипролиферативный эффект TGF-β, тогда как при астме недостаточность этих лигандов обеспечивает объединяющий механизм для повышенной эпителиальной восприимчивости и нарушения восстановления. . Это подтверждается обнаружением заметного снижения иммуноокрашивания эпителия для фосфотирозина (глобального маркера активации тирозинкиназы) в бронхиальном эпителии при легкой астме 40 и сообщениями об отсутствии активации MAPK 41.Напротив, при тяжелой астме, требующей кортикостероидов, активность тирозинкиназы заметно повышается в зависимости от тяжести заболевания 40.
При астме, хотя EGFR, c-erbB1, заметно сверхэкспрессируется, не наблюдается увеличения эпителиальной экспрессии связанных EGFR, HER-2 (c-erbB2) или HER-3 (c-erbB3) 40, тогда как при ХОБЛ и из этих рецепторов сверхэкспрессируются параллельно с EGFR (неопубликованные наблюдения). Поскольку гомодимеры EGFR: EGFR являются лишь слабыми активаторами пути MAPK 42, это будет дополнительно способствовать неспособности астматического эпителия противодействовать антипролиферативным сигналам, обеспечиваемым TGF-β.Напротив, гетеродимеры EGFR: HER2 являются мощными трансдукторами сигналов 43 и будут усиливать пролиферацию эпителия при ХОБЛ, в то время как гетеродимеры, содержащие c-erbB3, способны способствовать выживанию клеток благодаря наличию нескольких сайтов связывания для фосфатидилинозитол-3-киназы на этом рецепторе. 44 и родственный сигнальный субстрат для сигнальных рецепторов SMAD. Таким образом, продолжительность восстановления эпителия увеличивается при астме из-за дисбаланса между сигналами пролиферации и выживания, включающими семейство EGFR / HER, и антипролиферативными сигналами, включающими TGF-β.
Эпителиальная мезенхимальная трофическая единица
Компьютерная томография высокого разрешения, вскрытие и исследования биопсии при хронической астме выявили утолщение стенки дыхательных путей. Это включает отложение интерстициальных коллагенов в сетчатой пластинке 45, отложение матрикса в подслизистой оболочке, гиперплазию гладких мышц, а также микрососудистую и нейрональную пролиферацию. Утолщение ретикулярной пластинки является диагностическим признаком этого заболевания, и на основе измерений, проведенных в дыхательных путях человека 46 и в модели хронического воздействия антигена на морских свинках 47, 48, оно, по-видимому, отражает события, связанные с утолщением всей стенки дыхательных путей. 49.В 1990 году был описан слой субэпителиальных мезенхимных клеток с характеристиками миофибробластов, количество которых увеличивалось при астме пропорционально толщине 50 ретикулярного слоя коллагена. По расположению и функциям эти клетки соответствуют аттенуированной оболочке фибробластов, описанной Evans . и другие. 51 лежат рядом с lamina reticularis и формируют сеть, подобную звездчатым клеткам печени, которые, когда активируются повреждением печени, являются ключевыми эффекторными клетками, ответственными за фиброз 52.Поскольку бронхиальный эпителий находится в контакте с ослабленной оболочкой фибробластов, эти два клеточных слоя занимают ключевое положение для координации ответов на вызовы из вдыхаемой среды в более глубокие слои подслизистой оболочки 53.
Повреждение монослоев эпителия in vitro приводит к повышенному высвобождению фибропролиферативных и фиброгенных факторов роста, включая фактор роста фибробластов (FGF) -2, инсулиноподобный фактор роста (IGF) -l, фактор роста тромбоцитов (PDGF), эндотелин (ET) -1 и латентный и активный TGF-β 2 37, 54.Авторы также обнаружили, что TGF-β, FGF-2 и ET-1 увеличиваются при астме 55–57, причем TGF-β и FGF-2 зашифрованы во внеклеточном матриксе, как показано их совместной локализацией с декорином протеогликанов и гепарансульфатом. , соответственно 58. Для дальнейшего установления взаимосвязи между передачей сигналов EGFR в процессах репарации и ремоделирования авторы использовали EGFR-селективный ингибитор тирозинкиназы, который подавляет репарацию эпителия in vitro с результирующим увеличением высвобождения TGF-β 2 из поврежденных эпителиальных клеток 26.Это указывает на параллельные пути, действующие в репарации эпителиальных клеток, некоторые из которых направляют реституцию и регулируются EGFR, тогда как другие контролируют продукцию профиброгенного фактора роста независимо от EGFR. Предполагается, что при астме нарушение пролиферации эпителия заставляет эпителий бронхов дольше оставаться в фенотипе восстановления, что приводит к повышенной секреции профиброгенных факторов роста.
Использование модели совместного культивирования свидетельствует о том, что повреждение эпителия вызывает активацию миофибробластов.Полиаргинин (суррогат основных белков эозинофилов), механическое повреждение или искажение сливающихся монослоев эпителиальных клеток, выращенных на коллагеновом геле, засеянном миофибробластами легких человека, приводит к усиленной пролиферации и повышенной экспрессии гена коллагена из-за комбинированного воздействия FGF-2, IGF -1, PDGF-BB, TGF-β и ET-l 37. При умеренной астме ингаляционные кортикостероиды снижают воспаление дыхательных путей и уровни IGF-1 и FGF-2, но с ограниченным улучшением BHR и лишь частичным влиянием на отложение коллагена в lamina reticularis и уровни TGF-β 59.
Поскольку кортикостероиды снижают подслизистые эозинофилы на ~ 80%, устойчиво высокий уровень TGF-β в жидкости бронхоальвеолярного лаважа (БАЛ), скорее всего, является следствием поврежденного и восстанавливающегося эпителия и связанного с ним обновления матрикса в дополнение к эозинофилам. Поскольку как эпителиальная экспрессия EGFR 26, так и продукция TGF-β невосприимчивы к кортикостероидам, комбинированные эффекты этих сигнальных путей на EMTU могут обеспечивать механизмы ремоделирования тканей и объясняют неполное разрешение функции легких с помощью ингаляционных кортикостероидов, наблюдаемое при хронической астме.
Связь между эпителием и оболочкой субэпителиальных фибробластов напоминает процессы, которые управляют физиологическим ремоделированием дыхательных путей во время эмбриогенеза, когда эпителий и мезенхима действуют как «трофические единицы», регулирующие рост и разветвление дыхательных путей 60. Следовательно, авторы предполагают, что EMTU повторно активируется при астме, чтобы управлять патологическим ремоделированием дыхательных путей 53. У субъектов с бессимптомной BHR продольные исследования показали, что у тех, кто прогрессирует до астмы, наблюдаются параллельные изменения в воспалении и ремоделировании 61.Утолщение ретикулярной пластинки в биоптатах бронхов у детей раннего возраста также присутствует за несколько лет до того, как астма станет клинически проявленной. 17. Во время развития легких рост эпителия и мезенхимы частично регулируется балансом передачи сигналов EGF и TGF-β, как предполагается. при хронической астме. Предполагается, что у восприимчивых людей факторы окружающей среды взаимодействуют с EMTU в раннем возрасте, чтобы инициировать структурные изменения в дыхательных путях, которые могут объяснять снижение функции легких, наблюдаемое у маленьких детей, которые подвержены раннему хрипу 62, и потерю кортикостероидов. чувствительность к исходной функции легких, наблюдаемая в исследованиях CAMP и START 14, 15.Это подтверждается совсем недавними исследованиями на нечеловеческих приматах, где периодическое воздействие озона в присутствии аллергена создает фенотип, напоминающий хроническую астму 31. Следовательно, предрасположенность бронхиального эпителия, по-видимому, либо предшествует, либо возникает параллельно с факторами, предрасполагающими к Th3-опосредованному воспалению и является требованием для создания микросреды, в которой воспаление становится стойким в дыхательных путях и происходит ремоделирование.
Распространение реакций ремоделирования эпителиальной мезенхимальной трофической единицей
Для изучения мезенхимальных клеток, которые участвуют в реакции ремоделирования при астме, были разработаны протоколы их роста на основе биопсии бронхов.Эти фибробласты могут быть легко выращены из биопсий слизистой оболочки при астме и отличаются от фибробластов нормальных дыхательных путей в принятии характеристик стволовых клеток за счет быстрой пролиферации в отсутствие экзогенных факторов роста и их способности преодолевать контактное ингибирование в присутствии TGF-β 1 или TGF-β 2 , позволяющий достичь более высокой плотности клеток 63. В этом отношении обработка TGF-β 1 in vitro также заставляет подслизистые фибробласты принимать фенотип миофибробластов, что подтверждается индукцией актина α-гладких мышц. (α-SMA), приобретение сократительного фенотипа и синтез интерстициальных (репарационных) коллагенов.Хотя взаимосвязь между активацией миофибробластов и лежащей в основе гладкой мышечной массой при астме еще не изучена, после воздействия аллергена было обнаружено увеличение БАЛ гладких мышц и митогена сосудов, ЕТ-1 57, а лечение астмы с помощью TGF-β. фибробласты слизистой оболочки in vitro заставляли их высвобождать как ЕТ-1, так и другой митоген сосудов, фактор роста эндотелия сосудов (VEGF) 63. TGF-β приводит к значительному усилению продукции фактора роста соединительной ткани, и именно это, вероятно, является движущей силой большей части новое матричное производство.Помимо увеличения отложения матрикса, активация миофибробластов способствует пролиферации гладких мышц и микрососудов, которые являются характерными чертами реконструированных астматических дыхательных путей 64.
На кроличьей модели частичной обструкции оттока в мочевом пузыре TGF-β вызывает утолщение серозной оболочки из-за накопления миофибробластов, которые со временем изменяют фенотип на гладкомышечные клетки 65. В развивающихся капиллярах эндотелиальные клетки и гладкомышечные клетки также разделяют общий предшественник стволовых клеток с VEGF и PDGF, действующими как ключевые детерминанты клеточной судьбы 66.Исследования авторов нормальных и астматических фибробластов дыхательных путей (мио) показывают, что эти клетки проявляют фенотипическую пластичность, имеют некоторые ранние черты гладкой мускулатуры , например. белка SM-22 и может далее дифференцироваться с помощью TGF-β 1 и TGF-β 2 для экспрессии миозина тяжелой цепи и α-SMA 63, 67. Это предполагает, что клетки обладают свойствами обоих миофибробласты и гладкомышечные клетки, которые, вероятно, происходят от общего предка. Таким образом, медиаторы, высвобождаемые восстанавливающимся эпителием бронхов, обеспечивают механизм активации миофибробластов для распространения и усиления ремоделирования дыхательных путей.Такой механизм хорошо согласуется с недавними открытиями гладких мышц дыхательных путей, культивируемых из дыхательных путей при астме 64.
Взаимодействие между интерлейкином-4 и -13 и эпителиальной мезенхимальной трофической единицей
Воспаление дыхательных путей Th3-типа является характерным признаком астмы независимо от атопии. С учетом исследований на трансгенных мышах, которые предположили, что экспрессия трансгена IL-13 (но не IL-4) в эпителии бронхов приводит к ремоделированию подслизистой оболочки 68, 69, роль IL-4 и IL-13 в астматических эпителиальных клетках и функция фибробластов была исследована.Экспрессия высокоаффинной цепи IL-13Rα 2 ограничивается эпителиальными клетками дыхательных путей и фибробластами и коэкспрессируется с IL-13Rα 1 и IL-4Rα 70. И IL-4, и IL-13 передают сигнал через канал . фактор транскрипции STAT-6 71, экспрессия которого, как было показано, является заметной в бронхиальном эпителии и далее увеличивается при тяжелой астме 72. Хотя было показано, что IL-13 способен индуцировать трансформацию миофибробластов, он менее эффективен, чем TGF-β. и равносилен Ил ‐ 4 63.Поскольку параллельные эксперименты показали, что IL-13 вызывает нечувствительное к кортикостероидам увеличение высвобождения TGF-β 2 из эпителиальных клеток бронхов, кажется вероятным, что IL-13-опосредованное ремоделирование подслизистой оболочки инициируется в основном через бронхиальный эпителий 63. Недавнее исследование отчет Zhu et al. 73 поддерживает этот вывод, поскольку и фиброз, и гиперплазия гладких мышц в дыхательных путях мышей, экспрессирующих специфический для бронхиального эпителия трансген IL-13, зависят от TGF-β. Однако в эпителиальных клетках человека IL-4 так же эффективен, как и IL-13, в стимулировании высвобождения TGF-β, что повышает вероятность важных видовых различий в экспрессии эпителиального IL-4 и рецептора IL-13.
Хотя эффекты ремоделирования IL-4 и IL-13 можно отнести к активации эпителия, эти цитокины также оказывают прямое провоспалительное действие как на эпителиальные клетки, так и на фибробласты. Культуры бронхиальных эпителиальных клеток реагируют на IL-4 и IL-13 повышенным фосфорилированием STAT-6, сопровождаемым усилением колониестимулирующего фактора гранулоцитов / макрофагов и выработкой IL-8, которая дополнительно усиливается ферментативно активными экстрактами клеща домашней пыли ( Dermatophagoides pteronyssinus , Der p) 72, 74.Также было обнаружено усиленное высвобождение эотаксина из астматических фибробластов 63, что может помочь объяснить накопление эозинофилов под сетчатой пластинкой при астме. Таким образом, взаимодействуя с EMTU, IL-4 и IL-13 способствуют хроническому воспалению и ремоделированию дыхательных путей.
Заключение
Парадигма астмы с Т-хелперами типа 2 объяснила связь атопии с этим заболеванием, но не дала хорошего объяснения хронизации и тяжести заболевания.Чтобы принять это во внимание, авторы предполагают, что необходима вторая серия событий, при которых дыхательные пути ведут себя как хроническая рана с производством факторов роста, действующих на мезенхимальные клетки. Эта активация эпителиальной мезенхимальной трофической единицы обеспечивает необходимый стимул для изменения структуры и функции стенки дыхательных путей и может объяснить снижение функции легких, наблюдаемое с течением времени при этом заболевании, а также резистентный к кортикостероидам компонент BHR. Этот новый взгляд на хроническую астму обеспечивает основу для разработки новых терапевтических стратегий для решения остаточной неудовлетворенной клинической потребности, как недавно было показано положительное влияние рекомбинантного фактора роста эпителия человека при язвенном коллите, другом хроническом воспалительном заболевании, затрагивающем эпитлий 75.
Список литературы
- ↵
Banks-Schlegel S, Busse WW, Wenzel SE. Отчет о семинаре NIH NHLBI по патофизиологии тяжелой астмы. J Allergy Clin Immunol 2000; 106: 1033–1042.
- ↵
Чанг К.Ф., Годар П. Тяжелая резистентная к терапии астма: отчет рабочей группы ERS. Eur Respir Rev 2000; 10: 1–101.
- ↵
Пирс Н., Пекканен Дж., Бизли Р.Сколько астмы действительно связано с атопией? Thorax 1999; 54: 268–272.
- ↵
Leckie MJ, Brinke AT, Khan J, et al. Влияние моноклональных антител, блокирующих интерлейкин-5, на эозинофилы, гиперреактивность дыхательных путей и позднюю астматическую реакцию. Ланцет 2000; 356: 2144–2148.
- ↵
Брайан С.А., Канабар В., Матти С., и др. Влияние рекомбинантного человеческого интерлейкина-12 на эозинофилы, гиперчувствительность дыхательных путей и позднюю астматическую реакцию.Ланцет 2000; 356: 2149–2153.
- ↵
Skadhuage LR, Christensen K, Kyvik KO, Sigsgaard T. Влияние генетики и окружающей среды на астму: популяционное исследование 11 688 датских пар близнецов. Eur Respir J 1999; 13: 8–14.
- ↵
Rahman I, MacNee W. Окислительный стресс и регулирование глутатиона при воспалении легких. Eur Respir J 2000; 16: 534–554.
- ↵
Holgate ST.Генетическое и экологическое взаимодействие при аллергии и астме. J Allergy Clin Immunol 1999; 104: 1139–1146.
- ↵
Папи А, Джонстон SL. Риновирусная инфекция индуцирует экспрессию собственной рецепторной молекулы межклеточной адгезии 1 (ICAM-1) через усиление транскрипции, опосредованной NF-kappaB. J Biol Chem 1999; 274: 9707–9720.
- ↵
Reddel HK, Jenkins CR, Marks GB, et al. Оптимальный контроль астмы, начиная с высоких доз ингаляционного будесонида. Eur Respir J 2000; 16: 226–235.
- ↵
Исследовательская группа Европейской сети по изучению тяжелой астмы (ENFUMOSA). Качество жизни при тяжелой астме. Am J Respir Crit Care Med 2000; 161: A923.
- ↵
Исследовательская группа Европейской сети по изучению тяжелой астмы (ENFUMOSA). Поперечное европейское многоцентровое исследование клинического фенотипа тяжелой хронической астмы ENFUMOSA.Eur Respir J 2003; (Под давлением).
- ↵
Lange P, Parner J, Vestbo J, Schnohr P, Jensen G. Последующее 15-летнее исследование вентиляционной функции у взрослых с астмой. N Engl J Med 1998; 339: 1194–1200.
- ↵
Исследовательская группа программы управления детской астмой. Отдаленные эффекты будесонида или недокромила у детей с астмой. N Engl J Med 2000; 343: 1054–1063.
- ↵
Pauwels RA, Pederson S, Busse WW, et al. Раннее вмешательство будесонида при персистирующей астме легкой степени: рандомизированное двойное слепое исследование. Ланцет 2003; 361: 1071–1076.
- ↵
Cokugras H, Akcakaya N, Seckin I., Camcioglu Y, Sarimurat N, Aksoy F. Ультраструктурное исследование образцов биопсии бронхов от детей с умеренной астмой. Thorax 2001; 56: 25–29.
- ↵
Похунек П., Рош В.Р., Тарзикова Дж., Курдман Дж., Уорнер Дж.Эозинофильное воспаление слизистой оболочки бронхов у детей с бронхиальной астмой. Eur Respir J 1997; 10: Suppl. 25, 160с.
- ↵
Харт Л.А., Кришнан В.Л., Адкок И.М., Барнс П.Дж., Чанг К.Ф. Активация и локализация фактора транскрипции, ядерного фактора-каппаВ, при астме. Am J Respir Crit Care Med 1998; 158: 1585–1592.
- ↵
Уилсон С.Дж., Леоне Б.А., Андерсон Д., Мэннинг А., Холгейт СТ.Иммуногистохимический анализ активации NF-κB и экспрессии ассоциированных цитокинов и молекул адгезии в моделях аллергического воспаления у человека. Дж. Патол, 1999; 189: 265–272.
- ↵
Demoly P, Basset-Seguin N, Chanez P, et al. Экспрессия протоонкогена c-fos в биоптатах бронхов астматиков. Am J Respir Cell Mol Biol 1992; 7: 128–133.
- ↵
Sampath D, Castro M, Look DC, Holtzman MJ.Конститутивная активация эпителиального преобразователя сигнала и активатора пути транскрипции (STAT) при астме. Дж. Клин Инвест, 1999; 103: 1353–1361.
- ↵
Bertorelli G, Bocchino V, Zhuo X, et al. Повышенная регуляция белка теплового шока 70 связана с экспрессией HLA – DR при бронхиальной астме. Эффекты ингаляционных глюкокортикоидов. Clin Exp Allergy 1998; 28: 551–560.
- ↵
Puddicombe SM, Torres-Lozano C, Richter A, et al. Повышенная экспрессия ингибитора циклин-зависимой киназы p21 (waf) в эпителии бронхов при астме. Am J Respir Cell Mol Biol 2003; 28: 61–68.
- ↵
Chung KF, Barnes PJ. Цитокины при астме. Thorax 1999; 54: 825–857.
- ↵
Ordonez C, Ferrando R, Hyde DM, Wang HH, Fahy JV. Десквамация эпителия при астме. Артефакт или патология ?. Am J Respir Crit Care Med 2000; 162: 2324–2329.
- ↵
Puddicombe SM, Polosa R, Richter A, et al. Участие рецептора эпидермального фактора роста в восстановлении эпителия при астме. FASEB J 2000; 14: 1362–1374.
- ↵
Lackie PM, Baker JE, Gunthert D, Holgate ST. Экспрессия изоформ CD44 повышена в эпителии дыхательных путей у пациентов, страдающих астмой. Am J Respir Cell Mol Biol 1997; 16: 14–22.
- ↵
Ward PA, Gibson PG, Fakes K.Индуцированные эозинофилы мокроты при оценке астмы и хронического кашля. Респирология 2000; 5: 51–57.
- ↵
Bucchieri F, Puddicombe SM, Lordan JL, et al. Эпителий бронхов, страдающих астмой, более подвержен апоптозу, вызванному окислителями. Am J Respir Cell Mol Biol 2002; 27: 179–185.
- ↵
Нел А.Е., Диас-Санчес Д., Ли Н. Роль твердых частиц в воспалении легких и астме: доказательства участия органических химикатов и окислительного стресса.Curr Opin Pulm Med 2001; 7: 20–26.
- ↵
Миллер Л.А., Хайд Д.М., Гершвин Л.Дж., и др. Влияние аэроаллергена клещей домашней пыли и загрязнителей воздуха в младенчестве. Сундук 2003; 123: 434с.
- ↵
Гамильтон Л.М., Торрес-Лозано С., Пуддикомб С.М., и др. Роль рецептора эпидермального фактора роста в поддержании нейтрофильного воспаления при тяжелой астме. Clin Exp Allergy 2003; 33: 233–240.
- ↵
Demoly P, Simony-Lafontaine J, Chanez P, et al. Клеточная пролиферация слизистой оболочки бронхов у астматиков и хронических бронхитов. Am J Respir Crit Care Med 1994; 150: 214–217.
- ↵
Jost M, Kari C, Rodeck U. Рецептор EGF — важный регулятор множества эпидермальных функций. Eur J Dermatol 2000; 10: 505–510.
- ↵
Барроу RE, Ван Ч.З., Эванс М.Дж., Херндон Д.Н.Факторы роста ускоряют восстановление эпителия трахеи овцы. Легкое 1993; 171: 335–344.
- ↵
Виньола AM, Chanez P, Chiappara G, et al. Трансформирующая экспрессия фактора роста-бета в биоптатах слизистой оболочки при астме и хроническом бронхите. Am J Respir Crit Care Med 1997; 156: 591–599.
- ↵
Zhang S, Smartt H, Holgate ST, Roche WR. Факторы роста, секретируемые эпителиальными клетками бронхов, контролируют пролиферацию миофибробластов: модель совместного культивирования in vitro и ремоделирования дыхательных путей при астме.Lab Invest 1999; 79: 395–405.
- ↵
Kretzschmar M, Doody J, Massague J. Противоположные сигнальные пути BMP и EGF сходятся на медиаторе семейства TGF-beta Smad1. Природа 1997; 389: 618–622.
- ↵
Polosa R, Puddicombe SM, Krishna MT, et al. Экспрессия рецепторов и лигандов c-erbB в бронхиальном эпителии пациентов, страдающих астмой. J Allergy Clin Immunol 2002; 109: 75–81.
- ↵
Hamilton LM, Kimber I, Dearman RJ, et al. Фосфорилирование белкового тирозина в нормальном и астматическом бронхиальном эпителии. Иммунология 2000; 101: Дополнение. 1, 82.
- ↵
Koch A, Ito K, Tomita K, et al. Фосфорилирование митоген-активированных протеинкиназ p44 ERK1 и p42 ERK2 в биоптатах бронхов от астматиков и здоровых субъектов — Эффект теофиллина.Am J Respir Crit Care Med 2000; 161: A742.
- ↵
Lenferink AE, Pinkas-Kramarski R, van de Poll ML, et al. Дифференциальная эндоцитарная маршрутизация гомо- и гетеродимерных тирозинкиназ ErbB обеспечивает превосходство в передаче сигналов гетеродимерам рецепторов. EMBO J 1998; 17: 3385–3397.
- ↵
Graus-Porta D, Beerli RR, Daly JM, Hynes NE. ErbB-2, предпочтительный партнер гетеродимеризации всех рецепторов ErbB, является медиатором латеральной передачи сигналов.EMBO J 1997; 16: 1647–1655.
- ↵
Fedi P, Pierce JH, di Fiore PP, Kraus MH. Эффективное связывание с фосфатидилинозитол-3-киназой, но не с гамма-фосфолипазой C или белком, активирующим GTPase, отличает передачу сигналов ErbB-3 от передачи сигналов других членов семейства ErbB / EGFR. Mol Cell Biol 1994; 14: 492–500.
- ↵
Roche WR, Бизли R, Уильямс JH, Holgate ST. Субэпителиальный фиброз бронхов у астматиков.Lancet 1989; i: 520–524.
- ↵
Boulet LP, Laviolette M, Turcotte H, et al. Бронхиальный субэпителиальный фиброз коррелирует с реакцией дыхательных путей на метахолин. Chest 1997; 112: 45–52.
- ↵
Toda M, Yoshida M, Nakano Y, Cheng G, Motojima S, Fukada T. Животная модель утолщения стенки дыхательных путей. J Allergy Clin lmmunol 1997; 99: S409.
- ↵
Toda M, Yoshida M, Nakano Y, Cheng G, Motojima S, Fukada T.Утолщение базальной пластинки в стенке дыхательных путей на животной модели. Журнал Allergy Clin Immunol 1998; 101: S149.
- ↵
Chetta A, Foresi A, Del Donno M, Bertorelli G, Pesci A, Olivieri D. Ремоделирование дыхательных путей является отличительной чертой астмы и связано с тяжестью заболевания. Chest 1997; 111: 852–857.
- ↵
Брюстер CE, Ховарт PH, Джуканович Р., Уилсон Дж., Холгейт ST, Рош WR. Миофибробласты и субэпителиальный фиброз при бронхиальной астме.Am J Respir Cell Mol Biol 1990; 3: 507–511.
- ↵
Эванс MJ, ван Винкль LS, Fanucchi MV, Plopper CG. Аттенуированная фибробластная оболочка эпителиально-мезенхимальной трофической единицы дыхательных путей. Am J Respir Cell Mol Biol 2000; 21: 655–657.
- ↵
Артур MJ, Mann DA, Iredale JP. Тканевые ингибиторы металлопротеиназ, звездчатых клеток печени и фиброза печени. J. Gastroenterol Hepatol 1998; 13: Suppl.13, S33 – S38.
- ↵
Holgate ST, Davies DE, Lackie PM, Wilson SI, Puddicombe SM, Lordan JL. Эпителиально-мезенхимальные взаимодействия в патогенезе астмы. J Allergy Clin Immunol 2000; 105: 193–204.
- ↵
Howat WH, Holgate ST, Lackie PM. Высвобождение и активация изоформы TGF-бета во время заживления раны эпителия бронхов in vitro . Am J Physiol Lung Cell Mol Physiol 2002; 282: L15 – L23.
- ↵
Редингтон А.Е., Мэдден Дж., Фрю А.Дж., и др. Трансформирующий фактор роста бета 1 при астме. Измерение в жидкости бронхоальвеолярного лаважа. Am J Respir Crit Care Med 1997; 156: 642–647.
-
Редингтон А.Е., Рош В.Р., Мэдден Дж., и др. Основной фактор роста фибробластов при астме: измерение в жидкости бронхоальвеолярного лаважа базально и после провокации аллергеном.J Allergy Clin Immunol 2001; 107: 384–387.
- ↵
Makker HK, Springall DR, Redington AE, et al. Уровни эндотелина в дыхательных путях при астме: влияние эндобронхиального гипертонического раствора солевого раствора. Clin Exp Allergy 1999; 29: 241–247.
- ↵
Redington AE, Roche WR, Holgate ST, Howarth PH. Совместная локализация иммунореактивного трансформирующего фактора роста-бета 1 и декорина в биоптатах бронхов от астматиков и здоровых субъектов.Дж. Патол 1998; 186: 410–415.
- ↵
Hoshino M, Nakamura Y, Sim JJ, et al. Вдыхаемый кортикостероид снижает ретикулярную пластинку базальной мембраны за счет модуляции экспрессии инсулиноподобного фактора роста (IGF) -1 при бронхиальной астме. Clin Exp Allergy 1998; 28: 568–577.
- ↵
Варбертон Д., Шварц М., Теффт Д., Флорес-Дельгадо Г., Андерсон К.Д., Кардосо В.В. Молекулярные основы морфогенеза легких.Mech Dev 2000; 92: 55–81.
- ↵
Laprise C, Laviolette M, Boutet M, Boulet LP. Бессимптомная гиперреактивность дыхательных путей: связь с воспалением и ремоделированием дыхательных путей. Eur Respir J 1999; 14: 63–73.
- ↵
Stick S. Вклад развития дыхательных путей в заболевание легких у детей и взрослых. Thorax 2000; 55: 587–594.
- ↵
Рихтер А., Пуддикомб С.М., Лордан Дж., и др. Вклад интерлейкина-4 и интерлейкина-13 в эпителиально-мезенхимальную трофическую единицу при астме. Am J Respir Cell Mol Biol 2001; 25: 385–391.
- ↵
Johnson PR, Roth M, Tamm M, et al. При астме увеличивается пролиферация гладкомышечных клеток дыхательных путей. Am J Respir Crit Care Med 2001; 164: 474–477.
- ↵
Рулофс М., Фэджиан Л., Пампинелла Ф, и др. Трансформирующий фактор роста бета1 Участие в превращении фибробластов в гладкомышечные клетки в серозной оболочке мочевого пузыря кролика.Histochem J 1998; 30: 393-404.
- ↵
Yamashita J, Itoh H, Hirashima M, et al. Flk1-позитивные клетки, полученные из эмбриональных стволовых клеток, служат в качестве предшественников сосудов. Природа 2000; 408: 92–96.
- ↵
Chaudhary N, Richter A, Collins JE, Roche WR, Davies DE, Holgate ST. Сравнение фенотипов астматических и неастматических (мио) фибробластов. Am J Respir Crit Care Med 2001; 163: A473.
- ↵
Zhu Z, Homer RJ, Wang Z, et al. Легочная экспрессия интерлейкина-13 вызывает воспаление, гиперсекрецию слизи, субэпителиальный фиброз, физиологические аномалии и продукцию эотаксина. Дж. Клин Инвест, 1999; 103: 779–788.
- ↵
Ранкин JA, Picarella DE, Geba GP, et al. Фенотипическая и физиологическая характеристика трансгенных мышей, экспрессирующих интерлейкин 4 в легких: лимфоцитарное и эозинофильное воспаление без гиперреактивности дыхательных путей.Proc Natl Acad Sci USA 1996; 93: 7821–7825.
- ↵
Konstantinidis AK, Lordan JL, Bucchieri F, et al. Экспрессия рецептора IL-13 α2 эпителием бронхов человека. Am J Respir Crit Care Med 2001; 163: A737.
- ↵
Nelms K, Huang H, Ryan J, Keegan A, Paul WE. Механизмы передачи сигналов рецептора интерлейкина-4 и их биологическое значение. Adv Exp Med Biol 1998; 452: 37–43.
- ↵
Муллингс Р.Э., Уилсон С.И., Джуканович Р., и др. Повышенная экспрессия STAT6 в бронхиальном эпителии у пациентов с тяжелой астмой. J Allergy Clin Immunol 2001; 108: 832–838.
- ↵
Zhu Z, Ma B, Zheng T, et al. IL-13-индуцированные хемокиновые ответы в легких: роль CCR2 в патогенезе IL-13-индуцированного воспаления и ремоделирования. J Immunol 2002; 168: 2953–2962.
- ↵
Lordan JL, Bucchieri F, Richter A, et al.