Если ребенка укусил клещ
Если ребенка укусил клещ
Если ребенка укусил клещ: что делать и чего не делать, чтобы избежать тяжелых последствий.
По статистике детей клещи кусают чаще, чем взрослых, а домашних животных (например, собак) — чаще, чем детей. Однако именно дети наиболее сильно страдают от заболеваний, которыми клещи могут инфицировать человека после укуса.
Болезней, переносчиками которых являются клещи, немало — около десятка. Но наиболее опасных (а именно — смертельно опасных) из них две: клещевой энцефалит и клещевой боррелиоз.
Клещевой энцефалит (иначе — весенне-летний клещевой менингоэнцефалит). Это вирусная инфекция, которая характеризуется сильной интоксикацией и поражением головного и спинного мозга. Нередко клещевой энцефалит заканчивается стойкими неврологическими и психиатрическими осложнениями, а иногда, увы — смертью. Вирус клещевого энцефалита может существовать и размножаться в организмах около 130 видов животных и птиц.
Клещевой боррелиоз (или иначе — болезнь Лайма, или — Лайм-боррелиоз). Это довольно тяжелое бактериальное инфекционное заболевание, вызванное несколькими видами специфических бактерий, которые передаются человеку через укус клеща. Клещевой боррелиоз — самое распространенное заболевание, которое можно подхватить после «встречи» с клещом. Насекомые «цепляют» возбудителей этого заболевания у оленей, собак, птиц, овец, коров и других представителей фауны, а затем благополучно «пересаживают» их человеку при укусе вместе со своей слюной. Из ранних симптомов клещевого боррелиоза самые очевидные: жар; слабость; сильная головная боль; характерная сыпь, которая именуется в медицинской среде «мигрирующая кольцевидная эритема».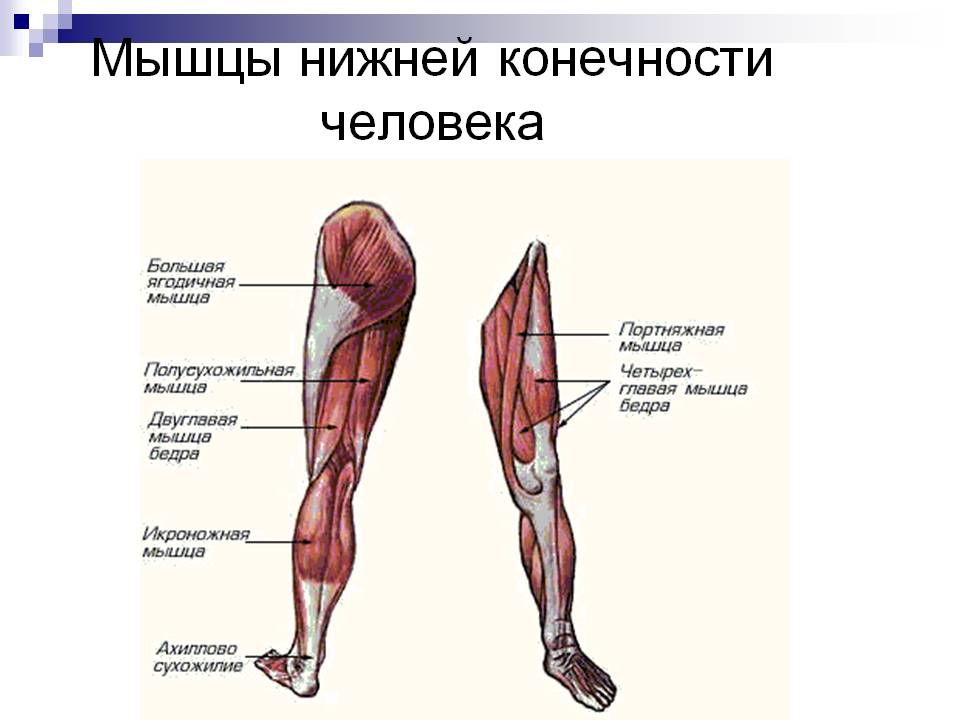
Куда чаще всего кусает клещ?
У клещей есть свои «любимые» места для укусов и присасывания. Причем, у детей и взрослых они разнятся — скорее всего. Например, у детей клещи чаще всего обнаруживаются на голове (и наиболее вероятно — за ушами), а у взрослых самое «популярное» место укусов — грудь, руки и подмышки. Кроме головы, клещи атакуют детей также в следующие области тела: шея и грудь; руки; подмышечная область (конкретно — у детей старше 10 лет).
Как удалить присосавшегося клеща в домашних условиях?
1. Тело клеща смазывают маслом и оставляют на 10-15 минут.
2. Затем необходимо сделать из плотной нити петлю и затянуть ее у основания хоботка клеща.
3. Придерживая кожу пальцами, покачивая клеща, постепенно вытягивать его, растягивая концы нити в разные стороны. Можно захватить клеща пинцетом или обернутыми чистой марлей пальцами, как можно ближе к его ротовому аппарату и, держа строго перпендикулярно поверхности укуса, повернуть тело клеща вокруг оси, извлечь его из кожных покровов.
4. Место укуса необходимо смазать 70% спиртом, 5% йодом, зеленкой или одеколоном.
5. Если клещ удален, а его головка осталась в коже, то нужно обратиться в поликлинику для лечебной помощи. Если обратиться нет возможности, то нужно обработать место 5% йодом и извлечь как занозу.
6. Клещей, снятых с тела, необходимо поместить в пузырек (в крайнем случае сжечь или залить кипятком). Не следует давить их пальцами, т. к. если клещ заражен, то вирус может попасть в организм человека через слизистые носа, глаз и незначительно поврежденную кожу.
7. После контакта с клещами обязательно вымыть руки с мылом.
8. Для определения зараженности клеща необходимо (в пузырьке, банке) доставить его в лабораторию. Клещей, снятых с человека, помещают в герметично закрывающийся контейнер с небольшим кусочком чуть влажной ваты. Контейнер помещают в холодильник при температуре +4С — +8 С (хранение в течение 1 суток), при длительном хранении необходимо заморозить при температуре – 20С (хранение в течение 2-х недель).
9. Если не удается извлечь самим, обратитесь в ближайшее медицинское учреждение.
Где провести исследование клеща на носительство инфекции?
Исследование снятых с людей клещей на зараженность возбудителями клещевого энцефалита, иксодового клещевого боррелиоза, и других возбудителей можно в следующих учреждениях:
|
Наименование филиала |
Адрес лаборатории |
Проводимые исследования |
|
ФБУЗ «Центр гигиены и эпидемиологии в Удмуртской Республике» |
г. Ижевск, ул. Ленина, 106 |
На наличие вируса клещевого энцефалита, боррелий, эрлихий, анаплазм и вируса ЛЗН |
|
Филиал ФБУЗ «Центр гигиены и эпидемиологии в Удмуртской Республике» в г. Глазове |
г. Глазов, ул. Кирова, 27а |
На наличие вируса клещевого энцефалита, боррелий |
|
Филиала ФБУЗ «Центр гигиены и эпидемиологии в Удмуртской Республике» в пос. |
Игринский район, п. Игра, ул. Труда, д. 17 |
На наличие вируса клещевого энцефалита, боррелий |
|
Филиал ФБУЗ «Центр гигиены и эпидемиологии в Удмуртской Республике» в г. Можге |
г. Можга, Ленина, 8 |
На наличие вируса клещевого энцефалита, боррелий |
|
Филиал ФБУЗ «Центр гигиены и эпидемиологии в Удмуртской Республике» в г. Сарапуле |
г. Сарапул, ул. Азина, 29 |
На наличие вируса клещевого энцефалита, боррелий |
|
Филиал ФБУЗ «Центр гигиены и эпидемиологии в Удмуртской Республике» в пос. Ува |
п. Ува, ул. Советская,7 |
На наличие вируса клещевого энцефалита, боррелий |
|
Филиал ФБУЗ «Центр гигиены и эпидемиологии в Удмуртской Республике» в г. |
г. Воткинск, ул. Свердлова,11 |
На наличие вируса клещевого энцефалита, боррелий |

 Воткинске»
Воткинске»
По состоянию на начало июня 2018 года в медицинские организации республики с жалобами на присасывание клещей обратились 7175 человек, из них – 1468 дети. Наибольшее количество обратившихся зарегистрировано в г. Ижевске (3519), в г. Можга (567), в г. Воткинске (501), в Увинском (195), Шарканском (204), Кезском (175), Игринском (148), Якшур – Бодьинском (141) районах. 4447 клещей исследованы на наличие возбудителя клещевого энцефалита, из них в 151 (3,4%) выявлен возбудитель клещевого энцефалита; 3387 — исследованы на наличие возбудителя клещевого боррелиоза, в т.ч. в 1240 клещах (36,6%) выявлен возбудитель иксодового клещевого боррелиоза.
По состоянию на начало июня 2018 года против клещевого энцефалита привито 45522 жителей республики, в том числе – 24956 детей в возрасте до 14 лет.
Если вашего ребенка укусил клещ, то бесплатно проводится иммунопрофилактика. Ее вы сможете получить в обслуживаемом медицинском учреждении (поликлиника) до 18.00. После 18.00 в стационарном учреждении по месту регистрации, для жителей г.Ижевска: БУЗ УР «ГКБ № 6 МЗ УР», БУЗ УР «ГКБ № 7 МЗ УР», БУЗ УР «ГБ № 3 МЗ УР».
Ее вы сможете получить в обслуживаемом медицинском учреждении (поликлиника) до 18.00. После 18.00 в стационарном учреждении по месту регистрации, для жителей г.Ижевска: БУЗ УР «ГКБ № 6 МЗ УР», БУЗ УР «ГКБ № 7 МЗ УР», БУЗ УР «ГБ № 3 МЗ УР».
Лучшая защита от клещей – это соблюдение техники безопасности:
1. Не рекомендуется без особой надобности залезать в непроходимые чащи низкорослого кустарника.
2. Перемещаясь по лесу, не срывайте веток, т.к. этим действием вы стряхиваете на себя клещей.
3. Ноги должны быть полностью прикрыты.
4. Спортивные трико, штаны должны быть заправлены в носки.
5. Обязательно наличие головного убора.
6. Длинные волосы желательно спрятать под головной убор.
7. После похода по лесу необходимо проверить и стряхнуть, как верхнюю одежду, так и нижнее белье.
8. Осмотреть все тело.
9. Тщательно расчесать волосы расческой с частыми зубчиками.
10. А лучшая профилактика от клещевого энцефалита – вакцинация. Вакцинацию проводят детям с 12 мес. возраста, для импортных вакцин, а с 4 лет — для отечественных.
А лучшая профилактика от клещевого энцефалита – вакцинация. Вакцинацию проводят детям с 12 мес. возраста, для импортных вакцин, а с 4 лет — для отечественных.
Стандартная схема вакцинации клещевого энцефалита состоит из 3 доз, которые вводятся по схеме:
— первая прививка осенью в выбранный день (октябрь, ноябрь)
— вторая прививка весной через 5-7 месяцев от первой,
— третья прививка через 9-12 месяцев от второй,
ревакцинация проводится каждые 3 года.
Для большинства вакцин разработана экстренная схема вакцинации. Целью применения экстренной схемы является быстрое достижение защитного эффекта, в случаях, когда сроки стандартной вакцинации были упущены.
— первая прививка весной(март, апрель) в выбранный день,
— вторая прививка через месяц от первой,
— третья прививка через 9-12 месяцев от второй.
ревакцинация проводится каждые 3 года.
Подробную информацию можете узнать у участкового педиатра, имеются противопоказания.
Полевые заметки — Подробный разбор режима карьеры в FIFA 21
Уважаемые поклонники FIFA!
Мы рады представить вам подробный разбор нововведений режима карьеры в FIFA 21!
В этом году мы хотели сделать особый акцент на режиме карьеры и предоставить вам в FIFA 21 игровые возможности, о введении которых вы просили. Год получился беспрецедентным, и нам всем пришлось преодолевать новые трудности, однако это не могло помешать нам представить на ваш суд режим карьеры, полный нововведений и новых игровых возможностей.
Нового действительно много, так что приступим.
Обзор
Основными задачами нашей работы над режимом карьеры было предоставить игрокам больше контроля, свободы выбора, глубины, разнообразия игры и сделать режим более реалистичным. Мы хотим, чтобы в матчах, трансферах и тренировках появилась новая глубина, и в то же время хотим сделать режим карьеры более доступным.
Интерактивная симуляция матчейМы знаем, что некоторые тренеры хотят контролировать все. Даже незначительные решения могут оказаться судьбоносными и определить исход момента, принести титул, привести к чемпионству. Однако не все моменты одинаково важны, поэтому в этом году мы хотели дать игрокам больше свободы в определении уровня контроля над своей командой.
Даже незначительные решения могут оказаться судьбоносными и определить исход момента, принести титул, привести к чемпионству. Однако не все моменты одинаково важны, поэтому в этом году мы хотели дать игрокам больше свободы в определении уровня контроля над своей командой.
Интерактивная симуляция матчей — это новый способ проведения встреч в режиме карьеры. Теперь вы можете отойти от традиционного игрового процесса и сосредоточиться на стратегии. Интерактивная симуляция матчей предоставляет более быстрый и тактический подход к матчам — тренер может принимать все важные решения из тренерской зоны и смотреть, как они воплощаются в жизнь, наблюдая за полем с высоты птичьего полета.
В интерактивной симуляции матчей игра проходит на удвоенной скорости. Неигровые моменты автоматически пропускаются, чтобы вы могли смотреть только на важные вещи.
Моментальный вход и выход из матча
В интерактивной симуляции матчей вы в любой момент можете вступать в игру и выходить из нее.
Вы можете брать управление в свои руки, когда ваша команда проигрывает, или вступать в игру всякий раз, когда игроки оказываются на ударной позиции, — интерактивная симуляция позволяет играть матчи именно так, как вам хочется. Кроме того, в любое время вы можете выйти из матча и посмотреть остаток симуляции или просто рассчитать финальный результат.
Как вход в игру и выход из нее, так и расчет финального результата можно сделать в любой момент матча, независимо от того, начали вы матч в обычном режиме или в режиме симуляции.
Новый способ игры в матче
Во время интерактивной симуляции матча вы увидите, как игроки вашей команды воплощают ваш план на игру, и получите все необходимые инструменты для внесения корректив прямо во время матча, если это потребуется. Вы можете использовать раздел «Управление командой», менять планы на игру и следить за показателями вашей команды и команды соперника в режиме реального времени. Вам доступны следующие вкладки:
- Физ.
 форма — здесь можно понять, у каких игроков вашей команды силы на исходе.
форма — здесь можно понять, у каких игроков вашей команды силы на исходе. - Оценки игроков — здесь видно, какие игроки проводят хороший матч, а у каких мало что получается.
- Статистика матча — моментальная сводка показателей игры обеих команд.
- План на игру — тактики, которые команды используют на поле.
Кроме того, в проводимых посредством интерактивной симуляции матчах будут присутствовать звуковые комментарии, а также атмосфера стадиона.
Новый запуск матчейВы можете начать матч, воспользовавшись одним из трех вариантов.
- Играть матч — загрузка обычного матча с традиционным игровым процессом.
- Симуляция матча — запуск новой интерактивной симуляции матча
- Расчет результата — моментальный расчет результат матча.
Чтобы помочь вам принять решение, мы сделали начало матчей более информативным. Теперь вы сможете увидеть вероятный состав соперника рядом с вашим текущим составом и сможете оценить важность матча и принять взвешенное решение.
Кроме того, мы постарались облегчить и ускорить процесс начала матча, внеся ряд изменений, например, автоматический выбор комплектов формы команд. Еще теперь здесь же размещены Параметры матча и некоторые новые настройки интерактивной симуляции матчей вроде возможности изменять отображение команд на схеме поля во время симуляции матча.
Развитие игроковРазвитие игроков и создание крепкого состава, способного привести вас к футбольной славе, — это неотъемлемая часть работы главного тренера, и мы знаем, что в прошлом комплектование состава в основном происходило за счет трансферов, а не за счет развития игроков. Теперь же с помощью новой игровой возможности «Развитие игроков» и полностью обновленной системой развития ваш состав сможет прогрессировать, и вы получите максимально возможный эффект от этого прогресса.
В зависимости от игровой позиции игроки смогут развиваться и специализироваться в определенном амплуа; кроме того, будут развиваться именно те характеристики игроков, которые соответствуют стилю вашей игры. Предпочитаете ли вы играть из глубины, много владеть мячом или проводить быстрые контратаки, вы сможете дать указания вашим игроками сосредоточить свое развитие в нужном вам направлении.
Предпочитаете ли вы играть из глубины, много владеть мячом или проводить быстрые контратаки, вы сможете дать указания вашим игроками сосредоточить свое развитие в нужном вам направлении.
Рост игроков определяется количеством набранного опыта. Количество получаемого опыта будет зависеть от того, насколько хорошо игроки проводят матчи (т. е. от игровой формы) и потенциала роста игрока. Полученный опыт распределяется по характеристикам и определяет развитие игрока в перспективе. По умолчанию все игроки используют сбалансированный план развития, что означает равномерное распределение опыта по всем характеристикам. Вы сможете назначить особый план развития игроку и выбрать определенные характеристики и особенности, на которые будет потрачен весь набранный опыт. Таким образом, вместо того, чтобы «разбазаривать» опыт на ненужные вам характеристики, план развития подразумевает распределение опыта на:
- основные характеристики (характеристики, от которых зависит ОБЩ рейтинг игрока на текущей позиции)
- Показатель особых приемов/игры нерабочей ногой (особенно для игроков атаки)
- Эффективность игрока в атаке/защите
- Характеристики, от которых зависит специализация (такие характеристики могут не влиять на ОБЩ на текущей позиции)
Как и в реальности, темпы развития игроков зависят от возраста, способностей, потенциала, формы и игрового времени. Дайте молодому игроку с большим потенциалом сыграть в нескольких матчах, помогите ему показать достойную игру, и он станет прогрессировать быстрее. Если же такого игрока надолго усадить на лавку, это даст обратный эффект — так что решать, что лучше для вашей команды, предстоит вам.
Дайте молодому игроку с большим потенциалом сыграть в нескольких матчах, помогите ему показать достойную игру, и он станет прогрессировать быстрее. Если же такого игрока надолго усадить на лавку, это даст обратный эффект — так что решать, что лучше для вашей команды, предстоит вам.
В ваших отзывах вы просили дать возможность тренировать игроков на других позициях. Теперь система развития игроков подразумевает возможность перевода игроков на новые позиции и предоставляет вам больше свободы и контроля над составом. Перевод на новую позицию может дать шанс игрокам, утратившим место в составе на своей изначальной позиции, таким образом, вы сможете применять более гибкий подход к каждому игроку.
Перевод на новую позицию использует аналогичную развитию игрока механику, однако ключевое отличие заключается в том, что весь набранный опыт направляется на характеристики и способности, отвечающие за игру на новой позиции.
Перевод на новую позицию может также повлиять на эффективность игрока в атаке или обороне и способность исполнять особые приемы. Например, вы хотите переквалифицировать вингера в крайнего защитника. Чтобы завершить перевод на эту позицию, нужно развить эффективность игры в обороне. Хотите перевести игрока ближе к атаке? Нужно улучшить исполнение особых приемов, чтобы игрок мог проходить защитников.
Например, вы хотите переквалифицировать вингера в крайнего защитника. Чтобы завершить перевод на эту позицию, нужно развить эффективность игры в обороне. Хотите перевести игрока ближе к атаке? Нужно улучшить исполнение особых приемов, чтобы игрок мог проходить защитников.
Молодым игрокам будет проще переквалифицироваться, а ветеранов можно будет перевести на позиции, где лучше сохранятся их ключевые характеристики.
Развитие и переквалификация игроков возможна и для воспитанников академии клуба. В академии молодые игроки постоянно находятся в хорошей форме и их развитие проходит равномерно.
В то время как планы развития используются для улучшения характеристик игроков в долгосрочной перспективе, новая система активных тренировок отвечает за общую готовность вашего состава к матчам посредством нового модификатора уровня игры футболиста — игрового тонуса. Подробнее об игровом тонусе мы поговорим ниже.
Система активных тренировок вводит понятие тренировочного дня, который может быть назначен на любой день за исключением дней проведения матчей.
- Каждый тренировочный день включает 3 отдельных тренировки, рассчитанные на определенные упражнения и определенных игроков.
- В каждой тренировке, в зависимости от ее типа, может принимать участие несколько игроков — всего в тренировочном дне может быть задействовано до 15 игроков.
- Каждая тренировка состоит из различных упражнений и отработки различных ситуаций, направленных на улучшение разнообразных аспектов игры: обороны, передач, дриблинга, ударов или стандартов.
- Тренировочный день всегда будет укомплектован подходящими тренировками и игроками из вашего текущего состава, однако вы можете самостоятельно редактировать и настраивать тренировочные дни.
- Смена текущего состава позволит вам моментально обновить выбор тренировок и игроков на тренировочный день — таким образом, вы сможете тренировать полностью новую группу игроков (например, ваших резервистов), разумеется, если у вас еще остались неиспользованные тренировки.
Давайте поговорим об этом подробнее:
- В зависимости от выбранного уровня сложности тренировочный день будет расходовать часть физической формы игроков.
 Чем труднее тренировочное упражнение, тем больше физической формы оно потребляет, однако и увеличение игрового тонуса будет существенным.
Чем труднее тренировочное упражнение, тем больше физической формы оно потребляет, однако и увеличение игрового тонуса будет существенным. - Максимального тонуса по результатам тренировки можно достичь, получив высшую оценку за проведенную или рассчитанную в симуляции тренировку. Расход физической формы будет неизменен вне зависимости от результатов тренировки.
- Разумеется, сложные тренировки требуют большего мастерства для получения высокой оценки. Тут у вас появляется выбор: действовать надежно и провести простую тренировку с гарантированно хорошей оценкой, чтобы получить максимум тонуса на этом уровне, или рискнуть, выбрать сложное упражнение и постараться получить высокую оценку и еще больше тонуса?
- Распределение тонуса зависит также и от количества игроков, участвующих в тренировке. Чем больше игроков участвуют в тренировке, тем больше игроков улучшит свой игровой тонус в результате успешного выполнения упражнения. Однако при меньшем количестве участников тренировки каждый игрок получит больше тонуса.

- Оценка, которую вы получаете за выполнение того или иного упражнения, зависит от количества набранных вами очков.
- При расчете упражнения в симуляции вы всегда будете получать высшую оценку, которую вы получили, сыграв в этом упражнении. По умолчанию при расчете результатов несыгранного вами тренировочного упражнения вы будете получать оценку D.
Тонус — это совершенно новый индикатор готовности игрока к матчу. У игрока с высоким тонусом повышаются основные характеристики, и он лучше играет в матчах.
Значение тонуса находится в диапазоне от 0 до 100.
- Тонус 50 означает, что у игрока средний игровой тонус. У такого игрока не будет ни усилений, ни штрафов к значениям основных характеристик. Таким образом, 50 — это базовое значение тонуса.
- Тонус в диапазоне 50-100 означает, что тонус игрока выше среднего и его характеристики будут усилены. Максимальный тонус (100) дает максимальную прибавку к характеристикам.

- Значение тонуса в диапазоне 0-50 означает, что тонус игрока ниже среднего и его характеристики будут понижены. Минимальный тонус (0) приведет к максимальному снижению характеристик.
Давайте рассмотрим действие тонуса на примере нападающего. Вот пример повышения характеристик игрока с тонусом 100:
- Выбор позиции +4
- Завершение +5
- Владение мячом +3
- Сила удара +3
Обратите внимание, что в данном материале приведены не окончательные значения ОБЩ и характеристик.
Если тонус достигнет ноля, навыки нападающего будут уменьшены:
- Выбор позиции -4
- Завершение -5
- Контроль мяча -3
- Сила удара -3
В управление командой была добавлена система индикации состояния игроков, выводящая на экран важную информацию об игроке и позволяющая принимать более взвешенные решения. Кроме возраста и развития игрока, теперь на его общий рейтинг влияют еще 3 элемента:
- Тонус
- Настрой
- Игра на своей позиции
Кроме того, рядом со значком настроя появился значок тонуса. Он поможет выделить игроков, находящихся в лучшей форме перед матчем.
Он поможет выделить игроков, находящихся в лучшей форме перед матчем.
Составление расписания позволяет контролировать действия игроков в каждый день сезона, чтобы поднимать их игровой тонус и управлять физической формой и настроем.
В долгосрочной перспективе баланс физической формы, настроя и тонуса имеет первостепенное значение, и этот процесс создает новое измерение управления игроками в режиме карьеры.
Теперь вы можете задать 3 типа ежедневной активности с четко разграниченными целями каждый:
- Тренировочный день — как описано в посвященном тренировкам разделе, основной упор тут сделан на повышение игрового тонуса за счет физ. формы и настроя (если тонус повышается слишком часто). Ухудшение физ. формы напрямую связано с тренировочной активностью, в то время как настрой… скажем так, ни один игрок не будет рад работать без выходных, и если он тренируется слишком часто, его настрой ухудшается.
- День восстановления — такие дни нужны для восстановления физической формы ваших игроков, однако во время восстановления тонус теряется быстрее всего.
- Дни отдыха — восстанавливается меньше физической формы, однако тонус понижается меньше, и немного улучшается настрой.
Составляя расписание, вы можете либо вносить изменения день ото дня, либо создать график на дни перед матчем, после матча и остальные дни между матчами на две недели вперед.
Если микроменеджмент не ваш конек, ничего страшного — графики можно корректировать в любой момент во время сезона, и вы сможете совершать ключевые изменения без необходимости планировать тренировки, отдых и восстановление игроков каждый день.
Календарь на месяц сохранит привычный функционал (просмотр следующих месяцев и расчет до нужной даты), но теперь будет также отражать заданную в расписании тренировочную активность. Здесь также можно корректировать графики на протяжении всего сезона. Если вы хотите вносить более глубокие изменения, перейдите в расписание на неделю, чтобы планировать действия на каждый день.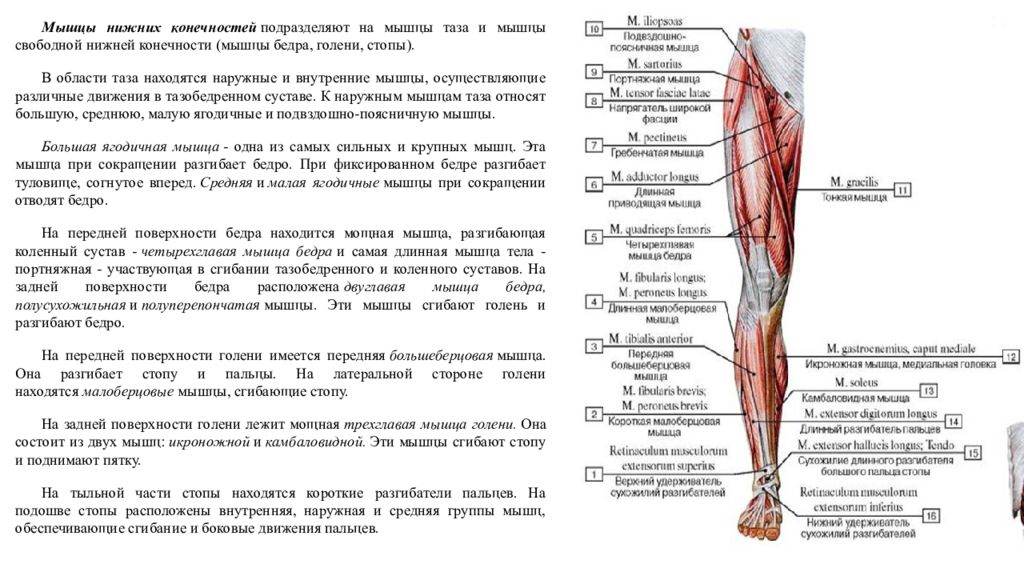
За последние годы современный трансферный рынок претерпел множество изменений, и мы хотели отразить это в игре. Вот 3 улучшения, сделанные в этой области:
- Аренда с правом выкупа — вместо совершения рискованного прямого трансфера теперь вы сначала можете арендовать желаемого игрока и выкупить его, когда и если он оправдает ваши ожидания.
- Предложения ИИ по обмену игроками — обмен игроков появился в режиме карьеры уже довольно давно, однако до сих пор ИИ предпочитал старые добрые трансферы игроков в обмен на деньги. Теперь ИИ будет делать предложения по обмену игроками. Когда вы захотите купить игрока, вам также может быть предложен обмен на одного из ваших игроков.
- Улучшения продления контрактов — клубы под управлением ИИ получат возможность продлевать контракты на основании рейтинга игроков в команде, и топ-игроки будут реже становиться свободными агентами. Разумеется, время от времени это будет случаться, так как лучшие игроки не всегда будут соглашаться на продление контракта.

Ознакомление
Мы постоянно вводим новые игровые возможности, поэтому донесение до игроков информации о том, как устроен режим карьеры, приобретает все более важное значение. В этом году мы делаем первый шаг в этом направлении и рассказываем игрокам о целях, которые преследует каждая система в игре, и как она помогает играть успешнее.
Расширенный пользовательский интерфейс в тематике турнира
В прошлых версиях игры мы последовательно вводили новые элементы интерфейса, оформляя его в тематике турнира, в котором принимает участие игрок. В FIFA 21 мы развили эту идею — теперь все экраны в режиме карьеры будут оформлены в стилистике турнира, в котором вы принимаете участие, для создания соответствующей атмосферы предстоящего матча.
Ниже приведены турниры, в стилистике которых будет оформляться интерфейс:
- Английская Премьер-лига (Англия)
- Лига 1 (Франция)
- Бундеслига (Германия)
- ЛаЛига (Испания)
- MLS (США и Канада)
- UEFA Champions League
- UEFA Europa League
- CONMEBOL Libertadores
- CONMEBOL Sudamericana
Влияние ответов на пресс-конференциях на настрой
Система влияния ответов на пресс-конференциях на всю команду была изменена. Теперь ваши ответы на послематчевых пресс-конференциях будут влиять только на настрой игроков, принимавших участие в матче. В предматчевых пресс-конференциях влияние будет оказываться только на настрой игроков, которые могут принять участие в матче.
Теперь ваши ответы на послематчевых пресс-конференциях будут влиять только на настрой игроков, принимавших участие в матче. В предматчевых пресс-конференциях влияние будет оказываться только на настрой игроков, которые могут принять участие в матче.
Настрой и форма игроков
В отзывах вы сообщали, что поддерживать хороший настрой всего состава слишком просто, поэтому мы добавили дополнительный уровень в систему влияния на настрой игроков. Теперь наиболее подходящий игроку тип ответов будет зависеть от оценки игрока в предыдущем матче и текущей формы игрока. Показавший невысокий уровень игры в предыдущем матче игрок получит больший бонус к настрою в случае выбора мотивирующего ответа, в то время как похвала окажет наибольшее влияние на игроков, показавших хорошую игру.
Готовая молодежная академия
Теперь в самом начале руководства клубом вам больше не придется самостоятельно создавать молодежную академию. В клубе уже будет работать молодежный скаут, а в составе академии будут проходить обучение несколько игроков. Их количество зависит от рейтинга Развития молодежи, кроме того, есть вероятность, что один из молодых игроков проявит потенциал звезды мирового класса.
Их количество зависит от рейтинга Развития молодежи, кроме того, есть вероятность, что один из молодых игроков проявит потенциал звезды мирового класса.
Талантливый игрок в вашей академии
Если вы оформите предзаказ* или купите издание Champions или Ultimate FIFA 21, то получите талантливого игрока в свою молодежную академию. При создании сохранения режима карьеры один игрок с большим потенциалом будет ждать вас в молодежной академии. Этот игрок будет иметь гражданство страны, которую представляет ваш клуб, и журналисты в своих новостных заметках, посвященных успехам этого игрока, будут упоминать, что это местный парень, который рос недалеко от клуба, за который теперь играет.
Чтобы проверить в начале режима Карьеры, имеете ли вы право на получение талантливого молодого игрока в академию, требуется подключение к сети. Сделав первоначальные настройки, вы сможете продолжить игру и без подключения к сети, как это всегда было.
Настройка приобретения команды новыми владельцами
При настройке нового сохранения режима карьеры у вас будет возможность задать бюджет, полученный командой в результате ее покупки новыми владельцами. Вы сможете дать команде круглую сумму в 500 миллионов на трансферы или начать почти ни с чем — выбирайте вариант, который вам по душе.
Вы сможете дать команде круглую сумму в 500 миллионов на трансферы или начать почти ни с чем — выбирайте вариант, который вам по душе.
Строгость переговоров
Некоторым из вас всегда нравилась возможность создать звездную команду без каких-либо ограничений, другие же считали, что такая свобода трансферов нарушает реалистичность и только портит впечатление от игры. Вместо того, чтобы выбрать один из этих подходов, мы решили предоставить вам возможность самостоятельно выбирать, как настроить свою карьеру.
- Строгие переговоры означают, что:
- Клубы не будут вести переговоров со своими главными соперниками или конкурентами.
- Игроки не будут заинтересованы в переходе в более слабую лигу или клуб, а также в клуб, с которым у них или их команды есть вражда.
- Игроки с талантом «Игрок одного клуба» не будут заинтересованы в смене клуба.
- Вы не сможете делать предложения игрокам, которые были подписаны клубом менее чем год назад.
Если установлено значение переговоров «Свободное», все приведенные выше ограничения не применяются, и вы можете совершать любые трансферы. Вам больше не придется проводить сезон, чтобы получить игроков, недавно совершивших трансфер в реальном футболе.
Вам больше не придется проводить сезон, чтобы получить игроков, недавно совершивших трансфер в реальном футболе.
Завершение карьеры игроками
Логика завершения карьеры игроком была изменена — теперь игрок будет объявлять о завершении карьеры только в последний год действующего контракта. Это позволит отодвинуть срок завершения карьеры у звездных игроков-ветеранов. Возраст завершения карьеры игроками также был изменен, став ближе к 40 годам.
Индивидуальность молодых и регенерированных игроков
Была исправлена ошибка, связанная с присвоением неверной индивидуальности и психологии молодых и регенерированных игроков. Из-за этой ошибки ответы на пресс-конференциях, пост-матчевых интервью в беседах с игроками оказывали некорректное влияние на настрой игроков.
Задачи руководства
Задачи руководства были изменены, их критерии стали понятнее, особенно в части популяризации бренда. Задачи, связанные с посещаемостью матчей и продажей абонементов были заменены на интересные и понятные болельщикам клуба этапы. Например, достижение определенного количества побед в сезоне, домашней победной серии или серии игр без поражений на выезде — и т.д. Кроме того, требование по предоставлению игрового времени молодым игрокам теперь распространяется на два сезона, чтобы у вас было больше возможностей его выполнить.
Например, достижение определенного количества побед в сезоне, домашней победной серии или серии игр без поражений на выезде — и т.д. Кроме того, требование по предоставлению игрового времени молодым игрокам теперь распространяется на два сезона, чтобы у вас было больше возможностей его выполнить.
Переговоры о трансферах и аренде
Мы обнаружили и устранили ряд неполадок, из-за которых значение решения игрока по трансферу во время трансферных переговоров получалось выше, чем было задумано. Вот некоторые факторы, влияющие на трансферные переговоры:
- Наличие или отсутствие отступных в контракте.
- Наличие или отсутствие других предложений по игроку или уже принятого предложения от другого клуба.
- Статус игрока — является ли игрок свободным агентом или внесен клубом в список трансферов или аренды.
- Совпадение страны, которую представляют оба клуба.
- Совпадение гражданства игрока и страны, которую представляет заинтересованный в приобретении игрока клуб.

- Общий рейтинг клуба-покупателя и рейтинг лиги, в которой он играет.
- Сравнение общего рейтинга игрока и общего рейтинга лиги клуба-покупателя.
- Сравнение продолжительности действующего контакта игрока и продолжительности предлагаемого контракта.
- Продолжительность аренды в случае аренды.
- Продолжительность нахождения игрока в текущем клубе. Чем больше времени игрок находится в клубе, тем выше вероятность перехода.
- Отличия роли игрока в текущем клубе от предлагаемой роли в новом клубе. Этот момент является очень важным, так как зачастую имеет большой вес при принятии решения о переходе. Игрок с большей вероятностью согласится на ключевую роль в новом клубе, если в текущем клубе он является игроком ротации.
- Наличие предпочитаемой позиции игрока в тактической схеме нового клуба. Например, ЦОП не захочет играть в клубе, играющем по схеме 4-4-2 без выраженной позиции опорника.
Вся эта информация сводится в итоговое значение, от которого зависит решение игрока по переходу или аренде. От этого счета будет также зависеть, какую позицию игрок займет при обсуждении зарплаты.
От этого счета будет также зависеть, какую позицию игрок займет при обсуждении зарплаты.
В оформление трансляций был внесен ряд изменений, направленных на улучшение реалистичности и впечатлений от игры. Вот некоторые из этих изменений:
- Полностью обновлена экранная графика оформления трансляций французских Лиги 1 Uber Eats и Лиги 2 BKT
- Знаменитая атмосфера — В FIFA 21 у множества клубов будут присутствовать собственные гимны по примеру YNWA «Ливерпуля» или гимнов «Манчестер Сити» или мадридского «Реала». Среди таких клубов:
- «Челси»
- «Боруссия Дортмунд»
- «Милан»
- «Атлетико Мадрид»
- Голевая музыка — музыка, воспроизводимая после гола на домашнем стадионе. Среди таких клубов:
- «Челси»
- «Боруссия Дортмунд»
- «Интер»
- Объявления на стадионах на языке соответствующей страны — Теперь игра на стадионах разных стран Европы станет еще реалистичнее, и вы сможете ощутить местный колорит благодаря объявлениям на стадионе на следующих языках:
- Французский
- Итальянский
- Немецкий
- Испанский
- Новые темы для обсуждения комментаторами во время матчей:
- ваш тренерский дебют
- обладатель награды лучшему тренеру месяца
- дебют перспективного молодого игрока
- достижение определенных отметок по сыгранным и выигранным матчам в качестве тренера
- возвращение в игру важного игрока, восстановившегося после травмы
- игра футболиста против своего бывшего клуба
- новые перфомансы для некоторых клубов, включая «Манчестер Сити».

- новый дизайн зрителей для некоторых клубов.
Благодарим вас за ваши отзывы, предложения и вашу поддержку на протяжении многих лет. Мы уверены, что режим карьеры в FIFA 21 получится крайне интересным и с нетерпением ждем ваших впечатлений.
Андрей Букур, Алекс Константинеску и команда разработчиков режима карьеры FIFA.
—
Другие инсайдерские материалы о FIFA читайте на странице «Полевых заметок».
Сезон FIFA 21 начнется 9 октября. Оформите предзаказ* или приобретите издание Champions или Ultimate FIFA 21 и получите воспитанника вашей академии в режим карьеры.
Примечание: Эта статья дает общее представление о том, над какими нововведениями режима карьеры работают разработчики в FIFA 21 для PlayStation 4, Xbox One и ПК. Мы постоянно ищем новые возможности для улучшения игры в FIFA, поэтому отдельные части данной статьи могут потерять актуальность при внесении изменений, направленных на то, чтобы каждый пользователь мог получать удовольствие от игры.
Уже можно оформить предзаказ* на FIFA 21 для PlayStation 4, Xbox One и ПК.
*Действуют условия и ограничения. Прочитайте подробности.
Обсуждайте все новости FIFA, будьте с нами в Facebook , читайте нас в Twitter и Instagram , в twitter наших разработчиков @EAFIFADirect, а также на официальных форумах FIFA. Подпишитесь на электронную рассылку и получайте информацию о продуктах, новостях, событиях и акциях EA SPORTS FIFA и EA.
Скарлатина: профилактика, симптомы и лечение
В детской поликлинике № 23 прошла лекция на тему: «Скарлатина у детей». Ее провела заведующая отделением Ксения Батрак. На лекции обсудили симптомы, лечение и профилактические меры.
Скарлатина — острое инфекционное заболевание, отличительной чертой которого является сочетание ангины и мелкоточечной сыпи на коже. Продолжительность инкубационного периода – от 1 до 10 суток.
Симптомы
· резкое повышение температуры,
· головная боль и общая слабость
· тахикардия,
· апатия и сонливость или, напротив, повышенная подвижность
· тошнота,
· боль в горле
· сыпь и другие.
Скрытый период скарлатины продолжается от 3 до 7 суток. Заболевание начинается остро с резкого нарушения самочувствия ребенка: он становится вялым, сонливым, жалуется на выраженную головную боль и озноб. Температура тела быстро достигает высоких цифр (38—40 °С в зависимости от степени тяжести болезни). Нередко в начальном периоде заболевания отмечаются тошнота и рвота. Однако следует отметить, что в наши дни скарлатина у детей, как и во взрослом возрасте, может протекать на фоне субфебрильной температуры до 37°С.
При скарлатине спустя несколько часов на коже у детей возникает специфическая сыпь в виде мелких ярко-розовых точек на покрасневшей коже. Сыпь более выражена на лице, боковых поверхностях туловища и в местах естественных кожных складок (паховых, подмышечных, ягодичных). Характерным признаком скарлатины является резкий контраст между ярко-красными «пылающими» щеками и бледным носогубным треугольником, на коже которого элементы сыпи отсутствуют. Внешний вид ребенка также привлекает внимание: помимо цветового контраста, лицо его одутловатое, глаза лихорадочно блестят.
Характерным признаком скарлатины является резкий контраст между ярко-красными «пылающими» щеками и бледным носогубным треугольником, на коже которого элементы сыпи отсутствуют. Внешний вид ребенка также привлекает внимание: помимо цветового контраста, лицо его одутловатое, глаза лихорадочно блестят.
Лечение
Чаще всего лечение проводят в домашних условиях. Больного ребенка изолируют. При тяжелом течении инфекции показана госпитализация на период не менее 10 суток. Затем в течение 12 дней ребенок должен находиться дома, его нельзя допускать в детский коллектив. Во время лечения ребенок должен соблюдать постельный режим и правильно питаться. Предпочтение отдается протертой легкоперевариваемой пище, приготовленной на пару или в отварном виде. Употреблять ее нужно небольшими порциями, не реже 4 раз в день. Врачи рекомендуют придерживаться лечебной диеты №13 по Певзнеру, а через две недели – диеты № 7. Также необходимо соблюдать питьевой режим.
Профилактика скарлатины
· избегать контакта с носителем заболевания,
· укреплять иммунную систему ребенка,
· соблюдать правила личной гигиены.
Если происходит вспышка заболевания в детском коллективе, накладывается карантин на неделю. В этот период ежедневно проводят осмотр взрослых и детей, контактировавших с больным. Детей, которые контактировали с больным скарлатиной, не пускают в детский сад или в первые два класса школы в течение 7 дней.
Если в семье есть заболевший скарлатиной ребенок, необходимо соблюдать следующие правила:
· ребенок должен находиться в отдельной комнате,
· следует ежедневно проводить влажную уборку с использованием раствора хлорамина,
· ухаживать за больным может только один член семьи, который должен минимизировать контакты с другими людьми,
· больной должен пользоваться отдельной посудой и предметами личной гигиены; его белье и одежду также нужно стирать отдельно, при этом использовать дезинфицирующие средства или кипятить, и проглаживать с обеих сторон.
· игрушки, с которыми играл ребенок, нужно тщательно вымыть с дезинфицирующим раствором и сполоснуть проточной водой. Мягкие игрушки необходимо постирать или вовсе избавиться от них.
Мягкие игрушки необходимо постирать или вовсе избавиться от них.
После выздоровления на протяжении месяца ребенок должен находиться под наблюдением врача. Затем проводят контрольный анализ крови и мочи. При отсутствии в организме патогенных бактерий, пациента снимают с диспансерного учета.
Дети, больные скарлатиной, не должны посещать детские учреждения.
При появлении симптомов, необходимо обратиться к врачу.
Адрес: Большой Сампсониевский пр., д. 62
Телефон: 596‑39-80
Сайт: https://dgp11.spb.ru/
Модель
Человеческой ноги предсказывает мышечные силы, состояния и энергию во время ходьбы
Abstract
Люди используют высокую степень избыточности при срабатывании суставов с различными комбинациями действий мышц и сухожилий, обеспечивающими одинаковый общий крутящий момент сустава. Как разрешение этих избыточностей, так и энергетика таких систем зависят от динамических свойств мышц и сухожилий, в частности, их отношения силы к длине. Современные модели ходьбы, в которых используются стандартные параметры при моделировании динамики мышцы-сухожилия, как правило, значительно переоценивают метаболическое потребление, возможно, потому, что они не учитывают адекватно роль эластичности.В качестве альтернативы мы утверждаем, что морфология мышечно-сухожилий ноги человека эволюционировала, чтобы максимизировать метаболическую эффективность ходьбы с самостоятельно выбранной скоростью. Мы используем подход, основанный на данных, чтобы оценить эту гипотезу, используя кинематические, кинетические, электромиографические (ЭМГ) и метаболические данные, полученные от пяти участников, идущих с самостоятельно выбранной скоростью. Кинематические и кинетические данные используются для оценки длины мышцы-сухожилия, мышечного момента рук и суставных моментов, в то время как данные ЭМГ используются для оценки мышечной активации.Для каждого испытуемого мы проводим оптимизацию, используя предписанную кинематику скелета, варьируя параметры, которые определяют кривую зависимости силы от длины каждого сухожилия, а также силу и оптимальную длину волокна каждой мышцы, одновременно стремясь минимизировать метаболические затраты и максимально согласовать с предполагаемыми данными.
Современные модели ходьбы, в которых используются стандартные параметры при моделировании динамики мышцы-сухожилия, как правило, значительно переоценивают метаболическое потребление, возможно, потому, что они не учитывают адекватно роль эластичности.В качестве альтернативы мы утверждаем, что морфология мышечно-сухожилий ноги человека эволюционировала, чтобы максимизировать метаболическую эффективность ходьбы с самостоятельно выбранной скоростью. Мы используем подход, основанный на данных, чтобы оценить эту гипотезу, используя кинематические, кинетические, электромиографические (ЭМГ) и метаболические данные, полученные от пяти участников, идущих с самостоятельно выбранной скоростью. Кинематические и кинетические данные используются для оценки длины мышцы-сухожилия, мышечного момента рук и суставных моментов, в то время как данные ЭМГ используются для оценки мышечной активации.Для каждого испытуемого мы проводим оптимизацию, используя предписанную кинематику скелета, варьируя параметры, которые определяют кривую зависимости силы от длины каждого сухожилия, а также силу и оптимальную длину волокна каждой мышцы, одновременно стремясь минимизировать метаболические затраты и максимально согласовать с предполагаемыми данными. совместные моменты. Мы обнаружили, что значения метаболических затрат на транспортировку (MCOT) наших участников могут быть правильно согласованы (в среднем 0,36 ± 0,02 предсказано, 0,35 ± 0,02 измерено) с приемлемой точностью крутящего момента сустава посредством применения единственного ограничения к метаболическому бюджету мышц.Соответствующие оптимальные наборы параметров мышцы-сухожилия позволяют нам оценивать силы и состояния отдельных мышц, устраняя избыточность при срабатывании суставов и предоставляя понимание потенциальных ролей и целей управления мышцами ноги на протяжении всего цикла походки.
совместные моменты. Мы обнаружили, что значения метаболических затрат на транспортировку (MCOT) наших участников могут быть правильно согласованы (в среднем 0,36 ± 0,02 предсказано, 0,35 ± 0,02 измерено) с приемлемой точностью крутящего момента сустава посредством применения единственного ограничения к метаболическому бюджету мышц.Соответствующие оптимальные наборы параметров мышцы-сухожилия позволяют нам оценивать силы и состояния отдельных мышц, устраняя избыточность при срабатывании суставов и предоставляя понимание потенциальных ролей и целей управления мышцами ноги на протяжении всего цикла походки.
Сведения об авторе
Нервно-мышечные системы часто используют избыточность при срабатывании суставов, при этом различные комбинации действий мышц и сухожилий создают одинаковый общий крутящий момент сустава. И разрешение этих избыточностей, и энергетика таких систем сильно зависят от соотношения силы и длины в мышцах и сухожилиях.Многие модели ходьбы человека не могут должным образом учесть эластичность из-за невозможности масштабирования параметров мышцы-сухожилия для разных людей, вместо этого полагаясь на исходные значения, взятые из исследований на трупах. Это может привести к неточным оценкам метаболического потребления, а также силы и состояния отдельных мышц. Вместо этого мы оцениваем параметры мышцы-сухожилия с помощью процедуры оптимизации на основе данных, проверяя гипотезу о том, что человеческая нога эволюционировала, чтобы максимизировать метаболическую эффективность ходьбы с самостоятельно выбранной скоростью.Мы обнаружили, что экспериментально наблюдаемое метаболическое потребление может быть согласовано с разумной точностью крутящего момента сустава путем добавления единственного ограничения на метаболический бюджет на каждую мышцу. Соответствующие наборы параметров мышцы-сухожилия использовались для вычисления оценки силы и состояния мышц, что позволило понять потенциальные роли и цели управления основными мышцами ноги на протяжении всего цикла походки.
Это может привести к неточным оценкам метаболического потребления, а также силы и состояния отдельных мышц. Вместо этого мы оцениваем параметры мышцы-сухожилия с помощью процедуры оптимизации на основе данных, проверяя гипотезу о том, что человеческая нога эволюционировала, чтобы максимизировать метаболическую эффективность ходьбы с самостоятельно выбранной скоростью.Мы обнаружили, что экспериментально наблюдаемое метаболическое потребление может быть согласовано с разумной точностью крутящего момента сустава путем добавления единственного ограничения на метаболический бюджет на каждую мышцу. Соответствующие наборы параметров мышцы-сухожилия использовались для вычисления оценки силы и состояния мышц, что позволило понять потенциальные роли и цели управления основными мышцами ноги на протяжении всего цикла походки.
Образец цитирования: Markowitz J, Herr H (2016) Модель ноги человека предсказывает мышечные силы, состояния и энергию во время ходьбы.PLoS Comput Biol 12 (5):
e1004912. https://doi.org/10.1371/journal.pcbi.1004912
https://doi.org/10.1371/journal.pcbi.1004912
Редактор: Адриан М. Хейт, Университет Джона Хопкинса, США
Поступила: 06.09.2015; Одобрена: 11 апреля 2016 г .; Опубликован: 13 мая 2016 г.
Авторские права: © 2016 Markowitz, Herr. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Наборы данных, используемые в этой публикации, можно найти по адресу: https://web.media.mit.edu/~hherr/markowitz_2016/.
Финансирование: Этот проект был поддержан Национальным управлением по аэронавтике и исследованию космического пространства (https://www.nasa.gov/) в рамках гранта № 6926843. Грант был получен HH. Спонсор не имел никакого отношения к дизайну исследования, сбору и анализу данных, принятию решения о публикации или подготовке рукописи.
Спонсор не имел никакого отношения к дизайну исследования, сбору и анализу данных, принятию решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Ходьба человека основана на сложном взаимодействии нескольких физиологических систем, каждая из которых демонстрирует некоторую степень избыточности. Нервная система управляет сокращением мышц, получая сигналы от множества различных нервных путей. Мышцы работают вместе, чтобы производить движение, но разные комбинации мышечных действий могут создавать одинаковый крутящий момент в данном суставе. Сухожилия обеспечивают связь между мышцей и костью, но энергия, передаваемая скелету, может поступать либо от активной мышцы, либо от податливости сухожилия.Понимание того, как люди разрешают эту избыточность, было давней проблемой в области нейробиологии и биомеханики [1, 2].
Знание того, как нервно-мышечная система распределяет нагрузку во время выполнения данной задачи, поможет понять цели контроля, которые определяют ее действия. Потенциальные цели (рассмотренные в [3]) включают планирование траектории суставов или минимизацию потребления метаболической энергии, активного мышечного объема или мышечной усталости. Однако без адекватного понимания ролей каждого компонента системы такие управляющие гипотезы остаются лишь предположениями.
Потенциальные цели (рассмотренные в [3]) включают планирование траектории суставов или минимизацию потребления метаболической энергии, активного мышечного объема или мышечной усталости. Однако без адекватного понимания ролей каждого компонента системы такие управляющие гипотезы остаются лишь предположениями.
Роль отдельных мышц и сухожилий в создании движения зависит от нервного импульса к мышцам и от свойств генерирования силы как мышцами, так и сухожилиями. Несколько режимов экспериментальных наблюдений дают представление об этих элементах. Электромиографию (ЭМГ) можно использовать для количественной оценки нервного возбуждения отдельных мышц, выявления того, какие мышцы вносят вклад в данное движение, и определения некоторой степени интенсивности [4–6]. Однако он ограничен вариабельностью сигнала, артефактами измерений и неточным отображением физиологии и мышечной силы.Ультразвуковые датчики недавно использовались для визуализации отдельных движений некоторых дистальных мышц ног и сухожилий in vivo [7–13], но они применимы только для небольших мышц и ограниченных задач.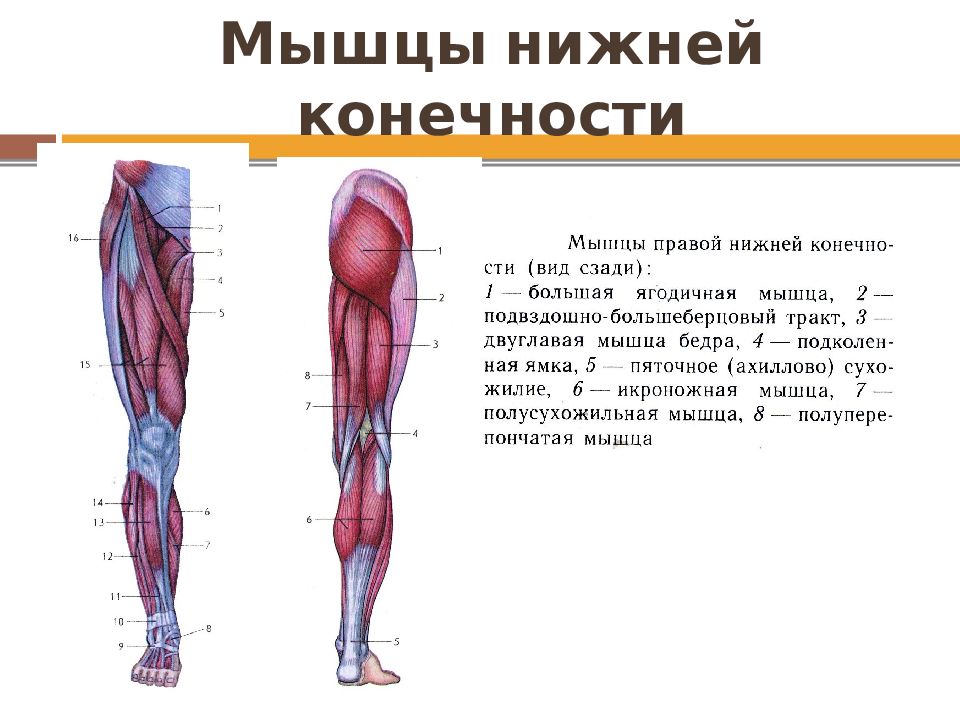 Захват движения может быть объединен со знанием анатомии, чтобы сделать вывод о чистом движении мышечно-сухожильных единиц при гораздо более общих обстоятельствах [14]; однако разделение результирующих профилей движения на отдельные вклады мышц и сухожилий требует знания часто недоступных параметров генерации силы в мышцах и сухожилиях.Эти параметры обычно оцениваются с помощью исследований на трупах, но масштабирование соответствующих количеств между различными мышцами и испытуемыми (не говоря уже о различиях с живыми образцами) недостаточно изучено [14–16] и может иметь значительное влияние на результаты моделирования. динамика [15, 16].
Захват движения может быть объединен со знанием анатомии, чтобы сделать вывод о чистом движении мышечно-сухожильных единиц при гораздо более общих обстоятельствах [14]; однако разделение результирующих профилей движения на отдельные вклады мышц и сухожилий требует знания часто недоступных параметров генерации силы в мышцах и сухожилиях.Эти параметры обычно оцениваются с помощью исследований на трупах, но масштабирование соответствующих количеств между различными мышцами и испытуемыми (не говоря уже о различиях с живыми образцами) недостаточно изучено [14–16] и может иметь значительное влияние на результаты моделирования. динамика [15, 16].
С учетом неполного представления, которое дают текущие экспериментальные измерения, желательна объединяющая теоретическая основа, которая объединяет доступные режимы данных в модель нервно-мышечной функции.Для решения этой проблемы при ходьбе были приняты два основных подхода: оптимальное управление и оптимальный дизайн.
Исследования ходьбы человека, основанные на оптимальном контроле [17, 18], моделируют морфологию сухожильных единиц (MTU) мышц ног с использованием оценок, основанных на литературе. Они определяют активацию мышц посредством оптимизации, выбирая цели управления, такие как минимизация метаболической энергии и / или отслеживание движения. Они были успешны в прогнозировании суставных моментов, суставных моментов и сил реакции опоры, но часто значительно переоценивали метаболические затраты, необходимые для передвижения [17, 19].Хотя очевидно, что нейронный контроль биологической системы каким-то образом оптимизирован, определение истинной целевой функции для этого подхода может оказаться невозможным. Множество различных комбинаций активации мышц могут давать одинаковые значения крутящего момента мышц, и сразу могут влиять несколько различных неизвестных целей управления и неврологических факторов. Кроме того, лежащая в основе неопределенность морфологии мышцы-сухожилия может привести к наблюдаемым избыточным метаболическим затратам, поскольку неправильное использование эластичности сухожилий может повлиять на силу и состояние мышц и, следовательно, на метаболические оценки.
Они определяют активацию мышц посредством оптимизации, выбирая цели управления, такие как минимизация метаболической энергии и / или отслеживание движения. Они были успешны в прогнозировании суставных моментов, суставных моментов и сил реакции опоры, но часто значительно переоценивали метаболические затраты, необходимые для передвижения [17, 19].Хотя очевидно, что нейронный контроль биологической системы каким-то образом оптимизирован, определение истинной целевой функции для этого подхода может оказаться невозможным. Множество различных комбинаций активации мышц могут давать одинаковые значения крутящего момента мышц, и сразу могут влиять несколько различных неизвестных целей управления и неврологических факторов. Кроме того, лежащая в основе неопределенность морфологии мышцы-сухожилия может привести к наблюдаемым избыточным метаболическим затратам, поскольку неправильное использование эластичности сухожилий может повлиять на силу и состояние мышц и, следовательно, на метаболические оценки.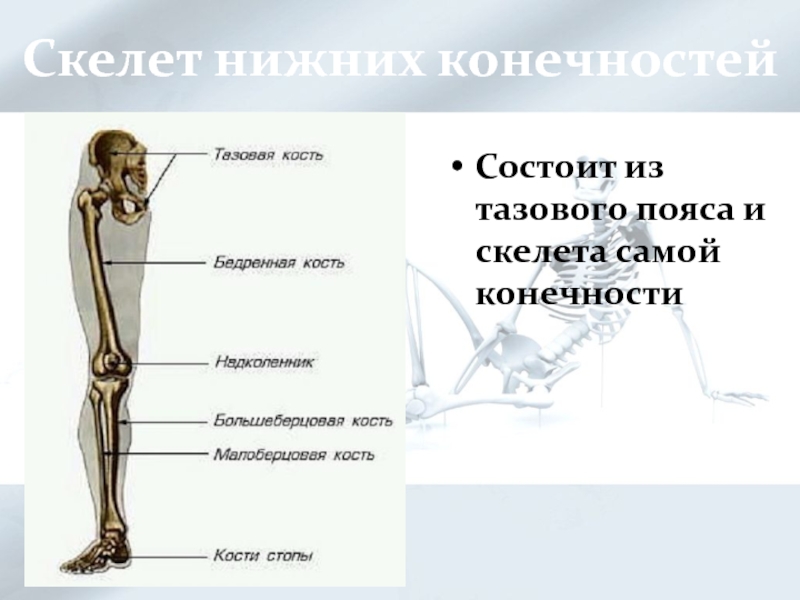
В моделях ходьбы человека, основанных на оптимальной конструкции, используется повышение эффективности, которое может быть достигнуто за счет настройки параметров MTU. Как показали Lichtwark и Wilson [20], экспериментально наблюдаемые деформации мышц и сухожилий можно предсказать, максимизируя эффективность изолированных MTU. Этот результат, вероятно, связан с хорошо задокументированной способностью сухожилия обеспечивать экономичную работу мышц [21, 22]. Кришнасвами и Герр [23] дополнительно исследовали потенциал оптимальной конструкции, оценивая разрыв крутящего момента мышц, охватывающих лодыжку, во время фазы опоры при ходьбе с самостоятельно выбранной скоростью.В этой работе использовались сигналы ЭМГ для оценки активации мышц во время ходьбы и была разработана схема оптимизации, основанная на предположении, что морфология мышечно-сухожильных единиц, охватывающих голеностопный сустав, эволюционировала, чтобы минимизировать метаболические затраты, необходимые для ходьбы с самостоятельно выбранной скоростью.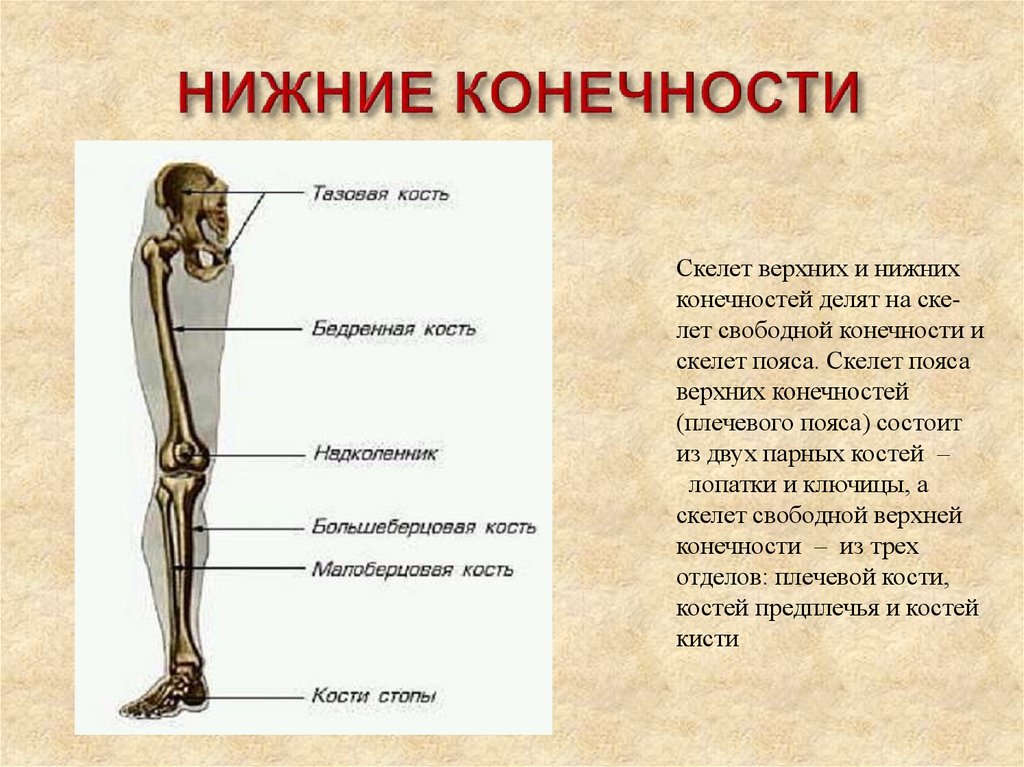 Его результаты показали, что один набор растворов может соответствовать как метаболическим, так и кинетическим параметрам человека, демонстрируя эффективное распределение нагрузки между подошвенными сгибателями, что качественно соответствует имеющимся экспериментальным данным.
Его результаты показали, что один набор растворов может соответствовать как метаболическим, так и кинетическим параметрам человека, демонстрируя эффективное распределение нагрузки между подошвенными сгибателями, что качественно соответствует имеющимся экспериментальным данным.
В этой работе мы продолжаем изучение [23], модифицируя и расширяя его, чтобы позволить исследовать всю ногу. Мы собрали кинематические, кинетические, электромиографические и метаболические данные от пяти субъектов, идущих с самостоятельно выбранной скоростью, и использовали их для оптимизации с заданной кинематикой скелета. Мы варьировали параметры, которые определяют, где каждая мышца действует на своей кривой «сила-длина», а также параметры, которые формируют соотношение «сила-длина» каждого сухожилия, стремясь одновременно минимизировать метаболические затраты и максимизировать согласие с наблюдаемыми суставными моментами.Каждая мышца в модели моделировалась как мышца типа Хилла [19, 24] и управлялась оценками активации, полученными на основе данных ЭМГ. Обзор модели системы и процедуры показаны на рисунках 1 и 2 соответственно. Мы обнаружили, что правильное метаболическое потребление может быть согласовано с разумной точностью моделируемого крутящего момента сустава за счет применения единственного ограничения на метаболический бюджет на каждую мышцу. Полученные оптимальные наборы параметров использовались для вычисления силы и состояния мышц, при этом профили длины пучка сравнивались с доступными экспериментальными измерениями.
Обзор модели системы и процедуры показаны на рисунках 1 и 2 соответственно. Мы обнаружили, что правильное метаболическое потребление может быть согласовано с разумной точностью моделируемого крутящего момента сустава за счет применения единственного ограничения на метаболический бюджет на каждую мышцу. Полученные оптимальные наборы параметров использовались для вычисления силы и состояния мышц, при этом профили длины пучка сравнивались с доступными экспериментальными измерениями.
Рис. 1. Модель системы.
Красные прямоугольники обозначают мышцы типа Хилла, а изогнутые линии обозначают податливые элементы. Все податливые элементы представляют собой связки, соединенные последовательно с мышцами, за исключением связки сгибателя бедра, которая обеспечивает пассивный момент сгибания в бедре.
https://doi.org/10.1371/journal.pcbi.1004912.g001
Рис. 2. Процедура идентификации мышечно-сухожильной системы.
Поле «Нелинейная динамика сокращения» включает в себя представления типа Хилла всех мышц в модели, а также нелинейную динамику сухожилий.
https://doi.org/10.1371/journal.pcbi.1004912.g002
Наши результаты организованы следующим образом. Сначала мы предоставляем наши предполагаемые профили активации мышц, обсуждая их качество и последствия. Во-вторых, мы отображаем результаты нашей двойной задачи оптимизации и резюмируем методологию, используемую для выбора одного оптимального решения для каждого предмета. В-третьих, мы оцениваем оптимальные решения, производя оценки энергетических переменных и состояния мышц. Мы сравниваем эти результаты с доступными экспериментальными измерениями, обнаруживая количественное согласие с метаболическими данными и качественное согласие с данными о длине мышечного пучка.
Результаты
Оценка активации мышц
Активация мышц обеспечивает масштабный коэффициент для способности данной мышцы генерировать активную силу в данный момент времени. Как описано в разделе «Методы», мы применили гибридный подход, аналогичный подходу [23], для оценки активации мышц по измерениям поверхностной ЭМГ пяти участников во время ходьбы.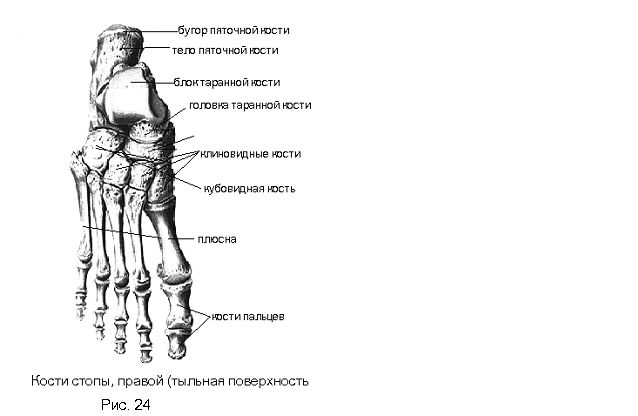 Байесовский алгоритм, впервые предложенный Сэнгером [25], был настроен (как описано в разделе «Методы») и использовался для выполнения оценки скрытого состояния, которая эффективно определяла нервное возбуждение каждой мышцы.Этот метод был выбран по сравнению с более традиционными методами полосовой фильтрации [4–6], потому что слегка запаздывающая синхронизация профилей, которые он производил, более легко позволяет получить наблюдаемые крутящие моменты в суставах. Мы более подробно остановимся на этом вопросе в разделе «Обсуждение». Предполагаемое нервное возбуждение затем передавалось на формирующий фильтр [26, 27], который представляет динамику активации мышц. Полученные средние профили представлены на рис. 3. Как видно из этого графика, произошли значительные различия (как внутри субъектов, так и между участниками) в оценках активации, полученных для мышц, охватывающих бедро.Это отсутствие согласованности, вероятно, было связано с некоторой комбинацией относительно большой глубины этих мышц под кожей, артефактов движения и относительной недоступности области.
Байесовский алгоритм, впервые предложенный Сэнгером [25], был настроен (как описано в разделе «Методы») и использовался для выполнения оценки скрытого состояния, которая эффективно определяла нервное возбуждение каждой мышцы.Этот метод был выбран по сравнению с более традиционными методами полосовой фильтрации [4–6], потому что слегка запаздывающая синхронизация профилей, которые он производил, более легко позволяет получить наблюдаемые крутящие моменты в суставах. Мы более подробно остановимся на этом вопросе в разделе «Обсуждение». Предполагаемое нервное возбуждение затем передавалось на формирующий фильтр [26, 27], который представляет динамику активации мышц. Полученные средние профили представлены на рис. 3. Как видно из этого графика, произошли значительные различия (как внутри субъектов, так и между участниками) в оценках активации, полученных для мышц, охватывающих бедро.Это отсутствие согласованности, вероятно, было связано с некоторой комбинацией относительно большой глубины этих мышц под кожей, артефактов движения и относительной недоступности области. Чтобы это не повлияло на последующий анализ, профили нервного возбуждения из экспериментов с проволочными электродами [28] были использованы для моносуставных мышц, охватывающих бедро. Эти профили нервного возбуждения были переданы в динамику активации из [26, 27] для согласованности во времени и впоследствии использованы в качестве входных данных для модели вместе с основанными на данных оценками активации для всех других мышц.Дополнительные сведения об этой процедуре приведены в разделах «Методы» и «Текст S1». Хотя нет доступной достоверной информации, с которой можно было бы сравнить наши оценки активации, мы отмечаем, что зависимость результатов от времени (при нормировании на процент цикла походки) была относительно неизменной для разных испытаний и испытуемых. Профили соответствуют ожидаемым временным шкалам наращивания и спада мышечной активации и, как видно из следующего, позволяют модели давать реалистичные кинетические и метаболические результаты.
Чтобы это не повлияло на последующий анализ, профили нервного возбуждения из экспериментов с проволочными электродами [28] были использованы для моносуставных мышц, охватывающих бедро. Эти профили нервного возбуждения были переданы в динамику активации из [26, 27] для согласованности во времени и впоследствии использованы в качестве входных данных для модели вместе с основанными на данных оценками активации для всех других мышц.Дополнительные сведения об этой процедуре приведены в разделах «Методы» и «Текст S1». Хотя нет доступной достоверной информации, с которой можно было бы сравнить наши оценки активации, мы отмечаем, что зависимость результатов от времени (при нормировании на процент цикла походки) была относительно неизменной для разных испытаний и испытуемых. Профили соответствуют ожидаемым временным шкалам наращивания и спада мышечной активации и, как видно из следующего, позволяют модели давать реалистичные кинетические и метаболические результаты.
Рис 3.Средние траектории активации для всех мышц всех испытуемых при скорости ходьбы, при которой метаболические затраты на транспортировку (MCOT) минимальны.
Каждый цвет представляет одного участника, а заштрихованные области представляют ± одно стандартное отклонение от среднего профиля.
https://doi.org/10.1371/journal.pcbi.1004912.g003
Идентификация параметра мышцы-сухожилия
Расчетная активация мышц a ( t ), а также кинематика сустава θ сустав ( t ), полученные из данных захвата движения, были использованы для приведения в действие полной модели ноги, показанной на рис. схема изображена на рис 2.Модель мышца-сухожилие ноги M была определена набором морфологических параметров, которые описывают характеристики генерации силы каждой MTU, а также двумя параметрами, которые позволяют генерировать пассивную силу подвздошно-бедренной, ишиофеморальной и пубофеморальной связками, а также другими соединительными связками. ткань на бедре. Эти связки, как известно, предотвращают чрезмерное растяжение бедра и позволяют восстанавливать упругую энергию в суставе [18, 29]; в частности, они снижают нагрузку на подвздошную мышцу вокруг пальца ноги. Вкладом каждого MTU были его максимальная изометрическая сила F max , общий масштабный коэффициент для длины провисания сухожилия l sl и оптимальная длина мышцы l opt , его эталонная деформация сухожилия λ ref , и коэффициент формы его сухожилия K sh . Сосредоточенная связка сгибателя бедра (HFL) действовала как простая поворотная пружина, параметризованная жесткостью пружины K HFL и углом зацепления θ 0, HFL .Каждая параметризация модели генерировала кинетические ( τ mod ( t )) и метаболические ( C ) выходные затраты:
(1)
Вкладом каждого MTU были его максимальная изометрическая сила F max , общий масштабный коэффициент для длины провисания сухожилия l sl и оптимальная длина мышцы l opt , его эталонная деформация сухожилия λ ref , и коэффициент формы его сухожилия K sh . Сосредоточенная связка сгибателя бедра (HFL) действовала как простая поворотная пружина, параметризованная жесткостью пружины K HFL и углом зацепления θ 0, HFL .Каждая параметризация модели генерировала кинетические ( τ mod ( t )) и метаболические ( C ) выходные затраты:
(1)
Вектор параметров варьировался с использованием схемы стохастической оптимизации с двумя целями [30], которая одновременно минимизировала метаболические затраты и разницу между вычисленными суставными моментами данных и моментами, созданными моделью. Границы были указаны, как описано в разделе «Материалы и методы»; они были выбраны достаточно широко, чтобы к ним не подходили выбранные оптимальные наборы параметров.Пространства решений для этой оптимизации показаны на рис. 4. Для каждого участника набор оптимальных по Парето решений (т. Е. Набор решений, при которых нужно было бы пойти на компромисс в отношении одной цели, чтобы улучшить другую) образует закругленный угол в цели. Космос. В идеальном случае этот угол должен быть острым и состоять из одного решения, оптимизирующего обе цели. Однако этот идеал обычно не встречается в шумных (реалистичных) системах и не встречается здесь.
Границы были указаны, как описано в разделе «Материалы и методы»; они были выбраны достаточно широко, чтобы к ним не подходили выбранные оптимальные наборы параметров.Пространства решений для этой оптимизации показаны на рис. 4. Для каждого участника набор оптимальных по Парето решений (т. Е. Набор решений, при которых нужно было бы пойти на компромисс в отношении одной цели, чтобы улучшить другую) образует закругленный угол в цели. Космос. В идеальном случае этот угол должен быть острым и состоять из одного решения, оптимизирующего обе цели. Однако этот идеал обычно не встречается в шумных (реалистичных) системах и не встречается здесь.
Рис. 4. Лучшее решение для каждого участника и его отношение к измеренным метаболическим затратам на транспортировку (MCOT; светло-серая полоса) и диапазон MCOT для всех участников (темно-серая полоса).
На этих графиках каждое пробное решение представлено точкой, значение x которой является требуемым MCOT, а значение y — средним R 2 его кинетических прогнозов по сравнению с измеренными моментами в голеностопном, коленном и тазобедренном суставах. .
.
https://doi.org/10.1371/journal.pcbi.1004912.g004
Для создания прогнозов на основе нашей модели мы выбрали один оптимальный набор параметров вдоль фронта Парето для каждого предмета. Это было достигнуто путем оценки метаболического потребления на каждую мышцу среди всех оптимальных по Парето решений (рис. 5).Было замечено, что в рамках набора решений те, в которых метаболические затраты были низкими, а кинетическая подгонка была плохой, имели стабильно низкие затраты на мышцу. Было замечено, что те, у кого были самые лучшие кинетические припадки, но относительно большие метаболические затраты, поглощали большое количество метаболической энергии в небольшом количестве мышц, что приводило к постепенному улучшению кинетической подгонки. Такое явление, вероятно, было вызвано шумом в данных (особенно в сигналах ЭМГ) и несовершенной способностью нашего сосредоточенного частичного набора мышц соответствовать силе, создаваемой всем набором человеческого тела. Нарастание метаболической энергии, наблюдаемое в этом подмножестве мышц, не является физическим, так как это привело бы либо к быстрой утомляемости, либо к моделируемой мышце гораздо большей, чем она есть на самом деле (поскольку мышечная масса масштабируется с F max ). Следовательно, мы исключили решения, которые демонстрировали такое поведение, выбрав в качестве оптимального оставшееся оптимальное по Парето решение с наилучшим кинетическим соответствием. Математически это было достигнуто за счет установки порогового значения дробного расходования широкой мышцы бедра, поскольку метаболические затраты этой группы мышц были самыми большими и значительно увеличивались по мере улучшения кинетической подгонки.Выбранное дробное отсечение позволило нам применить один критерий для каждого участника, чтобы соответствовать экспериментальным метаболическим затратам, как показано на рисунках 4 и 5. Таблица 1 показывает, как наш выбор оптимального решения может количественно соответствовать экспериментально наблюдаемым метаболическим затратам на транспортировку.
Нарастание метаболической энергии, наблюдаемое в этом подмножестве мышц, не является физическим, так как это привело бы либо к быстрой утомляемости, либо к моделируемой мышце гораздо большей, чем она есть на самом деле (поскольку мышечная масса масштабируется с F max ). Следовательно, мы исключили решения, которые демонстрировали такое поведение, выбрав в качестве оптимального оставшееся оптимальное по Парето решение с наилучшим кинетическим соответствием. Математически это было достигнуто за счет установки порогового значения дробного расходования широкой мышцы бедра, поскольку метаболические затраты этой группы мышц были самыми большими и значительно увеличивались по мере улучшения кинетической подгонки.Выбранное дробное отсечение позволило нам применить один критерий для каждого участника, чтобы соответствовать экспериментальным метаболическим затратам, как показано на рисунках 4 и 5. Таблица 1 показывает, как наш выбор оптимального решения может количественно соответствовать экспериментально наблюдаемым метаболическим затратам на транспортировку. по четырем из пяти предметов (и в среднем) при сохранении приемлемого соглашения о совместном моменте. Единственный субъект, у которого не было достигнуто количественного метаболического согласия, показал ошибку только 6%. Более подробная информация о сокращении включена в «Материалы и методы» и в S3 Text, а оптимальный набор параметров для каждого участника приведен в конце этого документа.
по четырем из пяти предметов (и в среднем) при сохранении приемлемого соглашения о совместном моменте. Единственный субъект, у которого не было достигнуто количественного метаболического согласия, показал ошибку только 6%. Более подробная информация о сокращении включена в «Материалы и методы» и в S3 Text, а оптимальный набор параметров для каждого участника приведен в конце этого документа.
Таблица 1. Эффективность многоцелевой оптимизации для выбранного решения для каждого участника, идущего с самостоятельно выбранной скоростью.
Показаны экспериментально измеренные и смоделированные метаболические затраты на транспортировку (MCOT), а также коэффициенты определения для смоделированных моментов голеностопного, коленного и тазобедренного суставов по сравнению с данными.
https://doi.org/10.1371/journal.pcbi.1004912.t001
Рис. 5. Подгонка полинома пятого порядка к дробным метаболическим затратам на мышцу как функции кинетического соответствия вдоль фронта Парето для каждого участника.
Черные линии представляют собой границы наблюдаемых метаболических затрат, а пунктирные синие линии представляют собой расположение выбранных оптимальных решений. Эти линии были получены путем сопоставления значений R 2 , перечисленных здесь на оси x, с соответствующими метаболическими затратами в решениях, показанных на рис. 4.
https://doi.org/10.1371/journal.pcbi.1004912.g005
Для дальнейшей оценки качества наших оценок момента суставов мы вычислили дробную среднюю абсолютную ошибку (FMAE)
(2)
между смоделированными суставными моментами τ mod и экспериментально наблюдаемыми суставными моментами τ obs .Эти величины вычисляются только в фазе опоры, чтобы облегчить сравнение с величинами, полученными из другого текущего анализа, основанного на ЭМГ, [31]. Величины, указанные в [31], представляют собой среднее значение по четырем различным видам деятельности (ходьба, бег, шаг в сторону и кроссовер), но в целом близки к тем, которые они генерируют только при ходьбе (за исключением бедра, у которого были более низкие ошибки при ходьбе). Их анализ включал два лечения; одна учитывала только одну степень свободы, а другая — несколько степеней свободы (как в нашей модели).В целом наш момент подходит для сравнения (таблица 2).
Их анализ включал два лечения; одна учитывала только одну степень свободы, а другая — несколько степеней свободы (как в нашей модели).В целом наш момент подходит для сравнения (таблица 2).
Обратите внимание, что здесь рассматривалась только фаза стойки. Цифры, приведенные в [31], представляют собой среднее значение по четырем различным видам деятельности, но в целом они близки к тем, которые они генерируют только для ходьбы. Их анализ включал два лечения; одна учитывала только одну степень свободы, а другая — несколько степеней свободы (как в нашей модели).В целом наш момент подходит к сравнению выгодно.
https://doi.org/10.1371/journal.pcbi.1004912.t002
Метаболические затраты нашей модели также согласуются с ранее опубликованными результатами. У всех испытуемых мы обнаружили, что средняя эффективность положительной мышечной работы составила 0,26 ± 0,02, что соответствует [23, 32, 33]. Мы также оценили метаболические затраты модели во время различных частей цикла ходьбы путем перекрестной ссылки на моделирование с данными входной силовой пластины.Результаты собраны в Таблице 3 и показывают разбивку, очень похожую на смоделированную в [34].
Таблица 3. Долевые затраты на метаболизм в фазах двойной опоры, одинарной опоры и качания цикла походки (если смотреть с одной ноги).
Видно, что среднее распределение для наших пяти участников приблизительно соответствует моделированию в [34].
https://doi.org/10.1371/journal.pcbi.1004912.t003
Поведение мышц в оптимальных решениях
Оптимальные наборы параметров мышцы-сухожилия, оцененные с помощью нашей процедуры оптимизации, предоставляют средства для устранения избыточности в совместном срабатывании для каждого субъекта.При применении в сочетании с рассчитанной кинематикой и расчетными активациями мышц можно оценить силу и состояние отдельных мышц. На рис. 6 показано распределение крутящего момента для каждого сустава в процентах от веса тела, умноженного на рост, и усредненное по всем испытуемым. На рис. 7 показаны траектории длины мышечного пучка, нормированные на l opt и усредненные по всем объектам. Обратите внимание, что разброс некоторых из этих профилей связан не с разницей в общей форме, а с общим смещением по длине, о чем свидетельствуют их почти постоянные стандартные отклонения.Длина камбаловидной мышцы, подколенного сухожилия и обширного пучка мышц на этом графике довольно хорошо согласуется как по форме, так и по смещению с предсказаниями [35]. Сила и длина камбаловидной мышцы также качественно согласуются с проекциями [11]. На рис. 8 показаны траектории скорости для каждого мышечного пучка, нормированные на его максимальное значение v max и снова усредненные по всем участникам. Как видно из небольшого разброса форм профилей на этих графиках, сила и состояние мышц следовали одинаковым траекториям у разных участников.Было также замечено, что эти предсказания мало меняются вдоль фронта Парето в экспериментально измеренном метаболическом диапазоне данного субъекта.
Рис. 6. Вклад отдельных мышц в крутящий момент суставов в виде процента веса тела, умноженного на рост, и усредненный по всем участникам.
Полосы ошибок представляют собой стандартное отклонение для разных участников.
https://doi.org/10.1371/journal.pcbi.1004912.g006
Рис. 7. Длины мышечных пучков, полученные с помощью модели, нормализованные по оптимальной длине мышцы и усредненные по всем испытуемым.
Полосы ошибок этих профилей представляют собой стандартное отклонение для разных участников, и их разброс в значительной степени обеспечивается постоянным смещением, а не различными формами, о чем свидетельствуют относительно постоянные стандартные отклонения (во времени) и рис. 8.
https://doi.org/10.1371/journal.pcbi.1004912.g007
Рис. 8. Скорости мышечных пучков, полученные с помощью модели, нормализованные по максимальным значениям ( v max ) и усредненные по всем участникам. .
Здесь положительная скорость относится к эксцентрическому движению (удлинению мышцы). Горизонтальные пунктирные линии при 0, -0,17, -0,30 v ce / v max представляют рабочие скорости мышц с низкой энергией, максимальной эффективностью и максимальной мощностью соответственно.
https://doi.org/10.1371/journal.pcbi.1004912.g008
В целом состояние мышечного пучка измерить чрезвычайно сложно. Единственным доступным в настоящее время способом получения этих профилей является ультразвуковое исследование, которое применимо только для относительно коротких дистальных мышц ноги.На рис. 9 смоделированные траектории мышц одного пациента сравниваются с экспериментальными профилями, доступными из опубликованных ультразвуковых исследований. В каждом случае длины мышечных пучков l м нормируются на их длину при ударе пятки, l mHS . Для построения этих графиков мы взяли смоделированную мышцу субъекта, который наиболее точно соответствовал среднему росту и весу в экспериментальном исследовании. Профили Soleus и gastrocnemius взяты из Ishikawa et al [8], профиль gastrocnemius взяты из Fukunaga et al [7], а профиль wideus lateralis взят из Chleboun [10].В подошвенных сгибателях длинные участки почти изометрической операции наблюдаются в средней стойке как в модели, так и в профилях in vivo и . В обширной мышце видно, что траектория пучка в некоторой степени отслеживает сгибание колена как в модели, так и в опубликованных данных. Однако, хотя качественные тенденции каждой мышцы согласуются, количественного согласия не наблюдается. Мы полагаем, что наблюдаемые различия связаны с (i) разницей в скорости ходьбы между нашим исследованием и литературой, (ii) естественными вариациями кинематики ранней стойки (что влияет на начальную длину мышцы для нормализации) и (iii) неопределенностью нарушение структуры мышц и сухожилий при ультразвуковых исследованиях.Мы надеемся, что будущие экспериментальные методики, возможно, с использованием имплантируемых датчиков, смогут дополнительно проверить наши предсказанные траектории пучков.
Рис. 9. Сравнение траекторий модельного пучка с данными ультразвукового исследования in vivo и [7, 8, 10].В каждом случае длины мышечных пучков l м нормированы на их длину при ударе пятки, l mHS . В нашем исследовании смоделированная мышца была взята у субъекта, который наиболее точно соответствовал среднему росту и весу в экспериментальном исследовании.
https://doi.org/10.1371/journal.pcbi.1004912.g009
Обсуждение
Можно сделать несколько наблюдений как относительно используемой методологии, так и результатов, полученных в этом исследовании. Что касается методологии, мы сначала рассмотрим шаги, предпринятые для оценки активации мышц в нашей модели. Как упоминалось выше, традиционный подход к оценке мышечного возбуждения на основе данных ЭМГ включает полосовой фильтр [4–6]. Мы попробовали этот подход с нашей схемой оптимизации (по всем предметам), но обнаружили, что он обеспечивает меньшую способность соответствовать наблюдаемым суставным моментам по сравнению с реализованным методом.Углубившись в это, мы обнаружили, что между подходами существуют два основных различия; (i) выбранный байесовский метод создает профиль, который включается и выключается более резко, чем сигнал, создаваемый полосовым фильтром, и (ii) сигнал, создаваемый байесовским методом, последовательно отстает от сигнала с пропускной полосой примерно на 50 мс. Резкость профиля не оказывает существенного влияния на результаты, так как в основном размывается последующей динамикой активации и усреднением. Однако зависимость от времени имеет значение; это отставание позволяет наращивать мышечную силу в соответствии с наблюдаемыми моментами в суставах.Интересно, что мы обнаружили, что [36] вводили задержку в 40 мс для их сигнала ЭМГ, чтобы «учесть электромеханическую задержку между поверхностной ЭМГ и производством силы». Эта задержка использовалась в других исследованиях [37, 38] и укладывалась в диапазон 10–100 мс, указанный для этих процессов в литературе [39, 40]. Мы обнаружили, что добавление задержки в 40 мс, как в [36], к возбуждениям, производимым ЭМГ-сигналами с полосовой фильтрацией, дало производительность почти так же хорошо, как и при использовании алгоритма Сэнгера, с меньшими вариациями между циклами походки.Однако этот метод с задержкой полосы пропускания не поддается такой биофизической интерпретации, как байесовская модель.
Несмотря на все наши усилия по сбору данных ЭМГ и оценке активации, недостатки в этой части нашего набора данных явно существуют. Поверхностная ЭМГ в целом подвержена шумам и артефактам, и алгоритм Сэнгера (проверенный только изометрически в его публикации [25]) не устраняет их. Хотя влияние несовместимых артефактов было минимизировано за счет исключения данных ЭМГ из явно скомпрометированных циклов походки и использования средних траекторий в нашей модели, они, вероятно, не были удалены полностью.Лучшие результаты могут быть получены в будущей работе с помощью тонкопроволочных измерений ЭМГ, которые, хотя и являются более инвазивными, как известно, дают более надежные сигналы. Здесь общие профили ЭМГ с тонкой проволокой, описанные в [28], были использованы для замены шумных поверхностных измерений ЭМГ мышц, охватывающих бедро. Интересно, что наша модель на самом деле показала немного лучшее согласие с наблюдаемыми профилями крутящего момента бедра, чем с профилями других суставов, но это вводит в заблуждение, поскольку большая часть смоделированного момента бедра исходила от подколенных сухожилий и связок сгибателей бедра (на которые не повлияли общие профили. ).
Более незначительным недостатком в трубопроводе ЭМГ была нормализация по значениям максимального произвольного сокращения (MVC). Хотя мы не знаем лучшей альтернативы, этот подход действительно привел к нормализованному возбуждению наших мышц, иногда превышающему единицу в тестах быстрой ходьбы. Когда это произошло, мы перенормировали значение в испытании быстрой ходьбы и обработали заново, но в других случаях константа нормализации могла быть слишком маленькой. К счастью, влияние этого масштабирования крайне минимально, поскольку предполагаемая активация напрямую умножает максимальную изометрическую силу F max , которая оптимизирована.Небольшой эффект остается, потому что нормализация происходит до того, как возбуждение претерпевает активационную динамику (5), но этот эффект в значительной степени не имеет отношения к нашим результатам.
Компонент идентификации системы в этом исследовании позволил разработать методологию оценки параметров мышцы-сухожилия, способную согласовать измеренное метаболическое потребление при создании профилей крутящего момента в суставах, которые достаточно хорошо отслеживали наблюдения. Один критерий, основанный на метаболическом потреблении группы обширных мышц (самой большой в модели), обеспечил метаболическое соответствие для всех участников.Хотя точность наших смоделированных профилей крутящего момента в суставах выгодно отличается от других текущих процедур моделирования, основанных на ЭМГ [31], они, как правило, недооценивают требуемые крутящие моменты в суставах. Эта характеристика, вероятно, связана с исключением некоторых мышц, а также скоплением некоторых групп мышц. В частности, дефицит голеностопного момента во время поздней стойки, вероятно, связан с исключением более мелких мышц-сгибателей подошвы, которые, как известно, задействуются в это время [28]. Точно так же образование комков в трех мышцах подколенного сухожилия (полуперепончатая, полусухожильная и длинная головка двуглавой мышцы бедра) может быть причиной некоторых недостатков, наблюдаемых в моментах сгибания колена на протяжении всего цикла походки.Дальнейшее устранение этих и других групп мышц, в которых активация мышц не совсем параллельна, а линии действия мышцы-сухожилия не совсем выровнены, вероятно, улучшит предсказательную силу этого подхода, позволяя более точно определять роли каждой мышцы во время данное задание.
Одним из примечательных аспектов модели является важность связки сгибателя бедра, которая обеспечивает почти весь необходимый момент сгибания бедра рядом с пальцем ноги без каких-либо метаболических затрат.Его линейная форма была выбрана, как в [29] для максимальной простоты, но не согласовывалась с нелинейной затухающей формой, использованной в предыдущей работе [18, 41]. Учитывая, что он произвел крутящий момент бесплатно, он мог подавить необходимое действие других сгибателей бедра (в частности, подвздошной кости) около пальца ноги и обеспечил лучшую посадку момента бедра, чем это было бы возможно при тех же метаболических затратах. Однако подавление других сгибателей бедра не могло быть значительным, поскольку эти мышцы недостаточно сильны для создания необходимого крутящего момента и не растягиваются значительно за этот период времени.Также известно, что тазобедренные связки создают момент сгибания, необходимый для уравновешивания верхней части тела против силы тяжести в положении стоя, когда линия силы тяжести проходит кзади от тазобедренного сустава [42]. Они позволяют людям стоять прямо и даже нести лишний вес без значительной работы мышц бедра, что позволяет поддерживать низкие метаболические затраты [43]. Поскольку использованные нами углы зацепления соответствуют стоянию, а демпфирование в соединительной ткани человека, как полагают, дает лишь небольшое отклонение от шкалы времени ходьбы [16], мы считаем, что вклад связки сгибателя бедра в нашу модель физиологически обоснован.
Оптимальные параметры мышц и сухожилий, найденные в этом исследовании, играют разные роли в обеспечении эффективного передвижения. Максимальная изометрическая сила мышц ( F max ) должна быть достаточно большой, чтобы соответствовать требованиям крутящего момента в каждом суставе, но достаточно маленькой, чтобы размер мышц и метаболические затраты были разумными. Длина провисания сухожилия l sl и оптимальная длина мышцы l opt определяют время создания силы, действуя согласованно с активацией мышц.Параметры формы сухожилия λ ref и K sh определяют упругие свойства сухожилия и настраиваются для обеспечения правильной работы мышц и срабатывания суставов. Полученные в результате мышцы и сухожилия вместе образуют взаимосвязанную систему, способную производить вращающий момент сустава, необходимый для передвижения, с минимальными метаболическими затратами.
Оценка скоростей, с которыми мышцы в нашей модели сокращаются во время активации, может пролить свет на цели их контроля.Как подчеркивалось в [29, 44], мышцы минимизируют метаболический расход на низких скоростях (т.е. когда действуют приблизительно изометрически). Далее А.В. Хилл [45] продемонстрировал, что скелетная мышца максимизирует свою эффективность при сокращении при v CE ≈ −0,17 v max и его выходной мощности при v CE ≈ −0,30 v макс . Эти три скорости ( v CE = 0, −0.17, -0,30 v max ) обозначены горизонтальными пунктирными линиями на рис. 8. Объединение этого с рис. 3 позволяет нам контекстуализировать моделируемые скорости каждой мышцы при активации. Видно, что в голеностопном суставе передняя большеберцовая мышца работает на низких скоростях. Это минимизирует метаболические затраты и согласуется с изометрическими сокращениями, смоделированными в [29, 44]. Видно, что как камбаловидная, так и икроножная мышца работают приблизительно изометрически за счет их активации в средней стойке перед быстрым увеличением скорости их сокращения в силовом ударе к концу стойки (≈ 60% GC).Требуемая положительная работа подошвенных сгибателей согласуется с необходимостью, отмеченной в [29, 44], но их быстрое сокращение в конце стойки не совсем соответствует цели эффективности камбаловидной мышцы и силовой цели икроножной мышцы, отмеченной в [ 23]. Это несоответствие составляет разность фаз; наблюдение в [23] было основано на скорости мышц при отведенном пальце ноги, но наши скорости подошвенных сгибателей достигают примерно тех же уровней около 53% GC. Видно, что в колене большинство мышц работают приблизительно изометрически.Это согласуется с [29, 44], в которых отмечается, что эти мышцы в первую очередь служат для регулирования жесткости сустава оптимальным и экономичным образом. Стоит отметить, что группа Вастуса повторно задействуется в конце замаха около режима оптимальной эффективности (разгибание колена для удара пяткой) перед возвращением в режим низкой скорости. Наблюдается, что большинство мышц бедра при активации сокращается с низкой скоростью. Двумя исключениями являются подвздошная мышца и длинная приводящая мышца, которые стремятся к максимальной эффективности, поскольку они сгибают бедро вокруг пальца ноги.Большая приводящая мышца также может приближаться к режиму максимальной эффективности, поскольку она расширяет бедро вокруг пятки, но это менее очевидно. Эндо и др. [29, 44] обнаружили, что мышцы, охватывающие бедро, нельзя моделировать строго изометрически, что согласуется с этими наблюдениями.
Существует несколько возможностей для расширения этой работы в будущих исследованиях. Один из способов — расширить модель за пределы сагиттальной плоскости; как показано в [31] согласование моментов в трех измерениях может повлиять на оптимизированные наборы параметров.Другой способ — проверить модель путем тестирования в различных условиях ходьбы. Если бы можно было получить более надежные измерения ЭМГ, мы могли бы обучить модель на основе ходьбы по ровной местности с самостоятельно выбранной скоростью, а затем оценить способность оптимального набора параметров соответствовать наблюдаемым профилям крутящего момента в суставах и метаболическим затратам в различных условиях (скорости , наклоны и т. д.) В этом случае новые экспериментальные наблюдения изменения функции подошвенного сгибателя в зависимости от скорости [12, 13] могут быть использованы для оценки модели.Другая возможность состоит в том, что наша общая гипотеза неполна — возможно, тело вместо того, чтобы адаптировать свою морфологию для максимальной эффективности при ходьбе (предположительно, наиболее распространенной задачи) с самостоятельно выбранной скоростью, на самом деле оптимизируется, имея в виду несколько задач. Можно представить себе дополнительный сбор данных от субъекта, бегущего с самостоятельно выбранной скоростью, и добавление дополнительных измерений к функции стоимости, в которой эта задача оптимизируется одновременно с ходьбой с самостоятельно выбранной скоростью. Таким образом, процедура, описанная здесь для самостоятельно выбранной скорости ходьбы, может стать частью более крупной схемы оптимизации за счет включения других задач.
Хотя такие эксперименты могут дать значительное понимание, они, несомненно, окажутся сложными из-за трудностей, связанных со сбором ЭМГ в указанных условиях. Одна из альтернатив — моделирование нейронного управления моделью. Недавние прямые динамические модели, основанные на рефлексивной обратной связи, показали способность стабильно ходить по разным ландшафтам [19, 46], с разной скоростью [47, 48] и в трех измерениях [48, 49]. Эти модели созданы для субъектов со «средними» размерами и, следовательно, могут быть улучшены за счет настройки на основе данных для отдельных субъектов и включения более реалистичных геометрий и морфологий мышц-сухожилий.Это может включать масштабирование инерции сегментов, уточнение линий действия мышцы-сухожилия и моментных рычагов [14], а также оптимизацию морфологии мышцы-сухожилия, чтобы одновременно минимизировать метаболические затраты и максимизировать согласие с экспериментально наблюдаемыми кинематическими и / или кинетическими данными. Таким образом можно получить более реалистичное представление об отдельном субъекте, что приведет к дальнейшему пониманию роли отдельных мышц и сухожилий во время походки и их различий между субъектами. Можно также представить себе изменение нейронного контроля в этой парадигме и использование структуры для изучения двигательных расстройств, таких как церебральный паралич.
Материалы и методы
Заявление об этике
Эксперименты данного исследования проводились в соответствии с принципами Хельсинкской декларации. Исследование было одобрено Комитетом Массачусетского технологического института по использованию людей в качестве субъектов эксперимента (протокол 1101004266). Перед началом экспериментов все участники дали письменное согласие на сбор, анализ и публикацию данных.
Сбор данных
Кинематические, кинетические, электромиографические и метаболические данные были собраны в лаборатории биологии скелета Гарвардского университета.В исследовании приняли участие пять здоровых взрослых мужчин со средним ростом, массой и возрастом 1,77 ± 5 м, 70,4 ± 6,3 кг и 26 ± 2 года соответственно. Необходимые наборы данных собирались в два этапа. Сначала испытуемые были оснащены переносной маской для потребления кислорода, прикрепленной к системе Cosmed K 4 B 2 VO 2 . В этой системе используется стандартный метод газового анализа открытого цикла для оценки потребления метаболической энергии на основе измерений вдыхаемого и выдыхаемого кислорода [50].Трем участникам было предложено постоять неподвижно в течение семи минут, пока регистрировали базальное измерение, при этом показатели двух других участников были приняты как среднее значение этих трех тесно сгруппированных показателей (1,38 ± 0,06 Вт / кг, 1,60 ± 0,07 Вт). / кг, 1,63 ± 0,06 Вт / кг). Затем участники ходили босиком по беговой дорожке с инструментами в течение семи минут на каждой из шести скоростей (0,75 м / с, 1,00 м / с, 1,25 м / с, 1,50 м / с, 1,75 м / с и 2,00 м / с), что позволило изменение расхода метаболической энергии, измеряемое в зависимости от скорости.Результаты были быстро сведены в таблицу и использованы для оценки скорости ходьбы при минимальных метаболических затратах на транспортировку (MCOT).
После того, как измерения метаболических затрат были завершены, маска потребления кислорода и система Cosmed были удалены, и каждый участник был экипирован для второй фазы. На этом этапе были собраны кинематические, кинетические и электромиографические данные для двух минут ходьбы босиком на каждой из семи скоростей; шесть из перечисленных выше и скорость, при которой MCOT объекта оказалась минимальной.Система инфракрасных камер (8 камер, Qualisys Motion Capture Systems, Гетеборг, Швеция) использовалась для отслеживания движения объектов, когда они шли в объеме захвата. Светоотражающие маркеры были размещены в 43 (двусторонних) местах на теле участника, и их трехмерные траектории были записаны с частотой 500 Гц. Расположение маркеров было выбрано специально для отслеживания движения суставов, как предписано моделью маркеров Хелен Хейз. Силы реакции опоры и контактные центры давления измерялись с использованием беговой дорожки с пластинами с разъемным ремнем (Bertec Corporation, Колумбус, Огайо).Электромиографические сигналы были собраны с использованием поверхностной системы от Motion Lab Systems (Батон-Руж, Лос-Анджелес) и размещения электродов, как предписано в [51]. Четырнадцать мышц (передняя большеберцовая мышца, камбаловидная мышца, медиальная икроножная мышца, широкая латеральная мышца, короткая мышца двуглавой мышцы бедра, прямая мышца бедра, полуперепончатая мышца, длинная головка двуглавой мышцы бедра, подвздошная мышца, большая ягодичная мышца (нижняя), большая ягодичная мышца (верхняя), средняя ягодичная мышца и длинная приводящая мышца magnus) на одной ноге каждого испытуемого, при этом симметрия предполагалась для другой ноги.Сигналы регистрировались на поверхности с помощью предварительно загущенных биполярных электродов (Electrode Store Model BS-24SAF, номер детали DDN-20), дискретизировались с частотой 1000 Гц и 20-кратно усиливались предварительными усилителями (Motion Lab Systems, номер детали MA- 411). Перед испытаниями при ходьбе для каждой группы мышц было проведено испытание максимального произвольного сокращения (MVC), в котором участника просили проработать эту конкретную группу как можно усерднее. Эти испытания MVC использовались для нормализации оценок мышечного возбуждения.
Оценка динамики суставов и геометрии мышц-сухожилий.
Данные маркера и силовой пластины были субдискретизированы до 125 Гц и обработаны с использованием SIMM (программное обеспечение для интерактивного моделирования опорно-двигательного аппарата, Musculographics Inc., Эванстон, Иллинойс). Статическое испытание использовалось для масштабирования динамической модели всего тела SIMM для каждого участника, при этом все последующие анализы основывались на этом масштабировании. Углы суставов, длина мышц-сухожилий и руки мышечно-сухожильного момента для каждого испытания были получены с помощью обратного кинематического анализа.Моменты суставов были рассчитаны с использованием SIMM Dynamics Pipeline, в котором используется программное обеспечение SDFAST (PTC, Needham, MA). Все полученные траектории были разбиты на циклы походки, временно нормализованы до процента цикла походки и усреднены. Грубые выбросы (обычно вызванные неправильной походкой или потерянными маркерами; порядка 5% циклов походки) были удалены перед усреднением.
Оценка активации мышц.
Активация мышц позволяет измерить способность мышцы генерировать активную силу в зависимости от времени.Он определяется как относительное количество кальция, связанного с тропонином в мышце, причем более высокие значения указывают на больший потенциал для образования поперечных мостиков и более высокие мышечные силы. Активация вызывается сигналами электрического нейронного возбуждения и поэтому может быть оценена с помощью измерений ЭМГ. Сигналы поверхностной ЭМГ представляют собой комбинацию потенциалов действия, токов деполяризации и ионных потоков внутри мышцы. Они предоставляют ценную информацию для определения вклада мышц, но из-за большой изменчивости сигнала и артефактов измерения должны быть тщательно обработаны, чтобы обеспечить надежные входные данные для количественных моделей.
Стандартный анализ ЭМГ моделирует сигнал как амплитудно-модулированный источник шума с ограниченной полосой [4], выпрямляя и фильтруя его нижними частотами для получения огибающей амплитуды [5, 6]. Несмотря на элегантность, этот подход не работал так хорошо в нашей задаче оптимизации, как недавно представленный Сенгером [25] метод. Как подробно описано выше, мы обнаружили, что небольшая задержка в профилях, созданных методом Сэнгера (по сравнению с методами, основанными на полосе пропускания), позволяет нашей модели более естественным образом наращивать требуемую мышечную силу.Метод Сэнгера моделирует сигнал ЭМГ как отфильтрованный случайный процесс со случайной скоростью и пытается вывести базовый нейронный управляющий сигнал x ( t ), который развивается как (3) В этом стохастическом дифференциальном уравнении с непрерывным временем dW — это дифференциал стандартного броуновского движения со скоростью α , dN β — это дифференциал процесса счета с β событиями, происходящими в единицу времени ( член скачка), а U — равномерно распределенная случайная величина в [0, 1].Без управляющего члена dN β это уравнение составило бы случайное блуждание. Скрытое состояние x ( t ) измеряется через (4) где P (emg | x ) — это условная вероятность наблюдения выпрямленного необработанного сигнала emg при заданном управляющем сигнале x ( t ). Эта модель отражает наблюдаемое лапласовское распределение соответствующих сигналов ЭМГ и объединяет все элементы измерения — различные волокна и электрические источники, фильтр кожи и жира, размещение и импедансы электродов — в одно уравнение.Комбинируя эти отношения с правилом Байеса и рекурсивно решая, Сэнгер получает максимальную апостериорную (MAP) оценку управляющего сигнала, то есть x ( t ), которая максимизирует P [ x ( t ). | emg ( т ), emg ( т −1), ..].
Результирующий управляющий сигнал, как было показано, точно отслеживает время включения и выключения профилей крутящего момента, создаваемых максимальными изометрическими сокращениями, при минимальном уровне шума, когда мышца неактивна [25].Однако, как отмечено в [23], он не может соответствовать ожидаемым сигналам активации, потому что (i) почти мгновенные переходы слишком резкие и (ii) профили не нарастают во время активации. Эти различия проистекают из несоответствия между смоделированным процессом прыжка-диффузии и истинным наращиванием мышечной активации. Внезапные скачки в [25] фиксируют начало и конец сигналов нервного возбуждения, но не учитывают время, необходимое для более медленной динамики связывания кальция. Кроме того, известно, что активное состояние мышцы нарастает быстрее, чем распадается [52].Эта функция отсутствует в модели Sanger; скорости диффузии и скачка симметричны относительно производной x ( t ). Комбинируя эти факты, становится ясно, что скрытое состояние x ( t ) более точно представляет нервное возбуждение мышцы, чем активное состояние мышцы. Для получения оценки активации применим динамику активации в виде [26, 27] (5) на выходе алгоритма Зангера x ( т ).Здесь a — активация мышцы, а τ act и τ deact — постоянные времени активации и деактивации, соответственно. Эти постоянные времени, как известно, зависят от состава мышечных волокон; конкретные значения, использованные для каждой мышцы в этом исследовании, приведены в тексте S2. Результирующая оценка объединяет время, полученное с помощью алгоритма Сэнгера, с известной биофизикой активации мышц.
Полный подход был реализован на собранных данных ЭМГ с использованием MATLAB (Mathworks, Natick, MA).Необработанный сигнал каждого канала данных ЭМГ был предварительно обработан путем удаления смещения постоянного тока, ограничения сигнала, превышающего 5 стандартных отклонений, нормализации до результирующего максимального значения и выпрямления. Отсечение было впервые включено Сэнгером, чтобы предотвратить попытки байесовского алгоритма оценить условные плотности по чрезвычайно редким значениям, что не может быть выполнено точно и, вероятно, является результатом артефактов. Затем данные были пропущены через байесовский алгоритм Сангера, описанный выше, для оценки нейронного возбуждения x ( t ) каждой мышцы (ссылка на исходный код в Справочнике).Члены диффузии и скачка ( α и β ) в [25] были скорректированы до 0,5 и 5 × 10 −31 , соответственно, чтобы обеспечить более надежный захват более слабых / более коротких сигналов без введения лишних случайных скачков в выходной сигнал или влияющий на его синхронизацию. Недостатком такой настройки было уменьшение резкости включения / выключения мышечных возбуждений, но это все равно произошло бы при последующей фильтрации и усреднении. Результирующие возбуждения были повторно нормализованы по значениям, оцененным во время испытаний на максимальное произвольное сокращение (MVC).Испытания MVC обрабатывались таким же образом, при этом в качестве значения MVC принималась средняя величина самого большого пакета (длительностью не менее 1 с). Иногда мы обнаруживали, что испытание MVC превосходило испытания быстрой ходьбы; в этом случае мы перенормировали на большее значение и повторно обработали. Затем для нормализованных профилей был установлен порог, чтобы удалить минимальный уровень шума, и они были переданы в формирующий фильтр (5), представляющий динамику активации мышц, которая была реализована с помощью решателя ode15s (жесткость / NDF) в MATLAB Simulink.Выходные профили активации и ( t ) были разбиты на циклы походки, временно нормализованы до процента цикла походки и усреднены. Грубые выбросы (около 10% циклов походки) снова были отброшены; в этом случае дополнительные источники ошибок включали артефакты движения и случаи плохого соединения электродов. Полученные в результате средние оценки активации нанесены на график со скоростью минимальной MCOT для всех субъектов на рис. 3.
К сожалению, не все каналы ЭМГ давали удовлетворительные оценки активации, при этом наблюдались низкие отношения сигнал / шум, особенно в мышцах, охватывающих бедро.Это было вызвано относительно сложной геометрией этого сустава, преобладанием связанных с ним артефактов движения, мышцами, расположенными глубоко под кожей, и относительным отсутствием доступа к этой области. Такие проблемы могут быть минимизированы за счет использования тонкопроволочных электродов, как в [28], и фактически мы обнаружили, что профили в [28] могут быть использованы для более точной оценки времени включения и выключения сигналов нервного возбуждения. для мышц, охватывающих бедро, чем наши измерения поверхности.Поэтому мы использовали эти литературные профили для получения профилей нервного возбуждения для подвздошной, длинной, большой и средней ягодичных мышц. Профили были оцифрованы с использованием MATLAB, в частности функции «ginput». Литературные профили даны в виде процентного цикла ходьбы; для каждого участника мы провели повторную выборку, чтобы продолжительность профиля соответствовала среднему циклу походки испытуемого. Никакой перенормировки амплитуды не потребовалось. Полученные профили затем пропускались через динамику (5) для получения оценок активации.В редких случаях, когда данные другой мышцы не могли быть восстановлены для данного субъекта, траектория этой мышцы принималась как среднее значение траекторий этой мышцы у всех субъектов, для которых измерение было приемлемым. Более подробная информация и статистический анализ этого подхода приведены в S1 Text.
Модель опорно-двигательного аппарата
Обработанные данные были использованы для построения динамической модели ноги при ходьбе (рис. 1). Модель включает в себя все мышцы, которые вносят значительный вклад в компоненты крутящего момента в голеностопном, коленном и тазобедренном суставах, перпендикулярном сагиттальной плоскости во время ходьбы.Несколько групп мышц были объединены для простоты; это считалось уместным, если все мышцы в группе имели схожие направления действия и активировались одновременно во время ходьбы. К ним относятся группа GAS (медиальная и латеральная икроножная мышца), группа VAS (латеральная широкая, медиальная и промежуточная мышца), группа HAM (полуперепончатая, полусухожильная и длинная головка двуглавой мышцы бедра) и группу ILL (подвздошная мышца и поясничная мышца). У каждой группы мышц было одно эффективное сухожилие, отражающее чистую податливость взаимодействия мышцы со скелетом.В дополнение к мышечно-сухожильным блокам была включена пассивная связка, охватывающая бедро, чтобы обеспечить восстановление упругой энергии в этом суставе. Этот элемент представляет вклад подвздошно-бедренной, седалищно-бедренной и лобково-бедренной связок, а также других соединительных тканей бедра и был смоделирован с использованием нелинейного уравнения в [18]; для простоты мы приняли это за линейную пружину, которая срабатывает только в то время, когда бедро становится вертикальным и начинает расширяться [29]: (6) Обратите внимание, что и момент, и угол в этом уравнении были определены с положительным сгибанием.Ниже мы описываем динамику системы мышц, сухожилий и суставов.
Мышечная динамика.
Все мышцы в модели имели динамику сокращения типа Хилла, аналогичную описанной в [19]. Каждая мышца содержит сократительный элемент (CE), который представляет активные мышечные волокна, и параллельный эластичный элемент (PE), который представляет эластичные структуры, окружающие мышцу. Сила сократительного элемента F CE зависит от активации мышцы a , длины сократительного элемента l CE и скорости сократительного элемента v CE .Сила параллельного упругого элемента F PE зависит только от длины сократительного элемента и действует только выше оптимальной длины волокна l opt . l opt и максимальная изометрическая сила F max зависят от размера мышцы, в то время как максимальная скорость сократительного элемента v max зависит от состава мышечных волокон.Для получения дополнительных сведений о реализованной модели мышц, включая предопределенные константы, см. S2 Text; подробности о том, как решались дифференциальные уравнения, см. в [19] и в соответствующем исходном коде.
Динамика сухожилий.
Как упоминалось выше, сухожилия — это нелинейные эластичные элементы, которые соединяют мышцы с костью. Их соотношение сила-деформация можно смоделировать в общем виде [16]: (7) куда (8) — деформация сухожилия за пределами его длины провисания l sl .Здесь F max — максимальная изометрическая сила мышцы, K sh — коэффициент формы, а λ ref — эталонная деформация. K sh определяет, где кривая «сила-длина» переходит от плоской нижней («носка») области к почти линейному поведению при больших деформациях. λ ref — деформация, при которой F SE = F max .Эти четыре параметра определяют морфологию конкретного сухожилия; F max и l sl шкала в зависимости от размера стержня, а K sh и λ ref зависят от свойств материала стержня.
Мышечно-сухожильная динамика.
Мышца и сухожилие действуют последовательно, но обычно ориентированы наклонно, причем угол между ними известен как угол перистости.Силы, действующие на сухожилие (SE), мышцу (M) и полный комплекс мышечных сухожилий, следующие: (9) Здесь длина сократительного элемента мышцы l CE представляет собой переменную состояния. Общая длина мышечно-сухожильного комплекса составляет (10) Угол перистости θ ( t ) изменяется таким образом, чтобы ширина мышцы оставалась приблизительно постоянной [15], и может быть записан как функция длины пучка: (11) Здесь угол θ 0 — это угол перистости, когда l CE = l opt (что очень близко к длине мышцы покоя [16]).Было обнаружено, что изменение этого угла в зависимости от длины пучка было необходимо для получения траекторий мышечных пучков, которые давали оценки метаболических затрат, аналогичные человеческим. Общий крутящий момент мышцы в данном суставе является суммой всех сил мышечно-сухожильных единиц, умноженных на их соответствующие изменяющиеся во времени моменты рычагов r i ( t ): (12) Чтобы устранить избыточность при срабатывании сустава, каждый раз необходимо определять изменяющуюся силу мышечно-сухожилия F MTC , i ( t ).
Оптимизация входов, выходов и параметров
Обзор примененной процедуры оптимизации показан на рис. 2. В нашей модели было две категории входных данных для каждой мышцы: (i) a ( t ), l mtc ( t ) и r i ( t ) — все оценены на основе данных и (ii) морфологических параметров мышцы-сухожилия, которые были определены с помощью оптимизации. Морфологические параметры, которые влияют на динамику сокращения смоделированных мышц по Хиллу, включают максимальную изометрическую силу мышцы F max , длину, при которой активная сила мышцы максимальна l opt , состав волокон FT , длина провисания сухожилия l sl , коэффициент формы сухожилия K sh и эталонная деформация сухожилия λ , ссылка .Мы оценили последствия изменения всех этих переменных, найдя разные уровни чувствительности для каждой. Переменные, к которым модель была нечувствительна, были фиксированы, оставив F max , K sh , λ ref и общий коэффициент масштабирования для l sl . и л выберите в качестве параметров для оптимизации. Последний фактор был выбран, чтобы гарантировать, что мышца работает в разумных областях своего пространства длины силы, предотвращая переобучение.Отношение l sl / l opt , как известно, значительно различается среди мышц, но не значительно в одной и той же мышце среди испытуемых [14], что оправдывает этот выбор.
Как упоминалось выше, наша оптимизация основана на гипотезе о том, что морфология мышцы-сухожилия мышц, составляющих ногу, эволюционировала, чтобы минимизировать метаболические затраты при ходьбе на скорости, при которой MCOT минимален. Это предположение подтверждается фундаментальной важностью двуногой ходьбы как средства передвижения и склонностью людей ходить с этой скоростью или приближаться к ней.Поэтому для каждого субъекта мы искали набор морфологических параметров, которые соответствуют измеренным крутящим моментам в суставах в голеностопном, коленном и бедренном суставах, потребляя при этом минимальное количество метаболической энергии. Это составляет проблему оптимизации с двумя целями, при которой функции метаболических и кинетических затрат одновременно минимизируются. Конкретные формы этих двух затрат кратко изложены ниже.
Оценка метаболических затрат.
Известно, что метаболическое потребление мышц зависит от нескольких факторов, включая размер пучка, уровень возбуждения, уровень активации, длину, скорость, силу и производство активной силы [24].Для данной мышцы в нашей модели все эти величины, за исключением уровней возбуждения и активации, зависят от морфологии мышцы-сухожилия и, следовательно, варьируются в зависимости от возможных решений. Для моделирования этих эффектов мы используем модель метаболического потребления, разработанную Умбергером и др. [24, 34]. Эта функция продемонстрировала исключительную прогностическую способность в ряде приложений и является наиболее широко распространенным из доступных показателей метаболических затрат. Он выражает метаболическую мощность на единицу мышечной массы как сумму четырех терминов: (13) Вот скорость нагрева активации, которая связана с переносом ионов Ca 2+ из саркоплазматической сети.- поддерживаемая тепловая мощность; и это, и скорость укорачивания / удлинения тепла обусловлены взаимодействием актомиозина. — скорость механической работы сократительного элемента, приведенная к мышечной массе. Полное математическое описание этих терминов см. В [24] и в дополнительных материалах к [34].
Чтобы оценить метаболические затраты всего тела, мы интегрируем каждую мышцу ноги, умножаем на каждую соответствующую мышечную массу и суммируем. Предполагалась двусторонняя симметрия, так как полные данные были собраны только для одной ноги каждого участника.Для каждой мышцы i масса M i была оценена с использованием (14) где ρ = 1059,7 кгм −3 и σ = 0,25 МПа — средняя плотность и удельное напряжение скелетных мышц, соответственно. Метаболизм остальной части тела (за исключением мышц ног) оценивали путем умножения оставшейся массы на измеренную базальную скорость ([Вт / кг]) стояния. Тогда окончательные метаболические затраты в модели C встретились с во временном окне T. (15) где M — общая масса тела, и суммы были взяты по всем мышцам обеих ног.
Кинетическая функция стоимости.
Помимо минимизации метаболических затрат, мы искали решения, которые наиболее точно воспроизводили измеренные моменты в суставах. Соответствующую стоимость C кин мы определили как (16) где значения R 2 — это коэффициенты определения между смоделированным и наблюдаемым крутящим моментом в шарнире.
Процедура идентификации системы
Как упоминалось выше, оптимальные параметры мышцы-сухожилия были определены с использованием двойной оптимизации.Для каждого субъекта было оптимизировано пятьдесят морфологических параметров; четыре морфологических параметра для каждой из двенадцати смоделированных мышц и два дополнительных параметра, описывающих связку сгибателя бедра. Выбранные границы параметров мышцы-сухожилия показаны в таблице 4. Максимальные значения изометрической силы ( F, , , , макс. ) для каждой мышцы должны были попадать в окно, окружающее масштабированное значение из SIMM. Большая ширина этого окна помогла компенсировать неопределенности в нормализации ЭМГ (через значения MVC, как обсуждалось выше).Коэффициент масштабирования для l слабины и l opt был выбран для обеспечения того, чтобы длина мышечного пучка оставалась в разумных физиологических рабочих диапазонах. Оценки для K sh и λ ref были взяты из [16]. Пределы упругости связки сгибателя бедра были выбраны таким образом, чтобы связка могла обеспечить от нуля до всего требуемый момент сгибания бедра вблизи пальца ноги.Угол зацепления был выбран таким образом, чтобы сухожилие могло включиться при сгибании бедра не более чем на 10 °, поскольку его физиологическая роль заключается в предотвращении чрезмерного растяжения.
Таблица 4. Границы параметров задачи оптимизации.
Значения максимальной изометрической силы ( F max ) для каждой мышцы были вынуждены попадать в довольно широкое окно, окружающее масштабированное значение из SIMM. Коэффициент масштабирования для l слабины и l opt был выбран для обеспечения того, чтобы длина каждого мышечного пучка оставалась в пределах физических рабочих диапазонов.Здесь параметр w определяет ширину отношения активной силы к длине мышцы; его значение зависит от мышцы, и все значения приведены в тексте S2. Оценки для K sh и λ ref были взяты из [16]. Пределы упругости связки сгибателя бедра были выбраны таким образом, чтобы связка могла обеспечить от нуля до всего требуемый момент сгибания бедра вблизи пальца ноги. Угол зацепления был выбран таким образом, чтобы сухожилие могло взаимодействовать с бедром, согнутым не более чем на 10 °, поскольку его физиологическая роль заключается в предотвращении чрезмерного растяжения.
https://doi.org/10.1371/journal.pcbi.1004912.t004
Модель была построена с использованием MATLAB и Simulink и интегрирована с использованием решателя ode15s (жесткое / NDF). Вычисления были распараллелены и выполнены с помощью Mathworks Cloud Center, компьютерного кластера, управляемого через Amazon Web Services. Этот ресурс оказался полезным, так как наши прогоны были настроены для тестирования многих решений и занимали около 10 часов на одном процессоре. В качестве алгоритма оптимизации использовался gamultiobj MATLAB, управляемый элитарный генетический алгоритм, который является вариантом алгоритма NSGA-II [30].Этот метод был выбран из-за вероятного присутствия локальных минимумов в обоих объективах и настроен, как показано в таблице 5. Оптимизационные настройки были выбраны для обеспечения достаточно больших поколений, чтобы заставить оптимизатор тщательно обыскивать пространство, устраняя необходимость в посеве населения. Каждому пробному раствору давали возможность работать в течение двух циклов походки, при этом учитывались только результаты второго цикла походки. Это устранило начальные переходные эффекты и позволило оценить систему в течение полного цикла походки.
Выбор оптимального решения.
Результатом нашей многоцелевой оптимизации является фронт Парето, набор решений, в которых одна функция затрат не может быть уменьшена без ущерба для другой. В идеальном случае этот фронт состоит из одного решения, в котором все цели оптимизированы; однако это редко происходит в шумных системах. Наша двойная оптимизация приводит к округленному фронту Парето, из которого мы выбрали одно «лучшее» решение.
Как видно на рис. 4, наши оптимальные по Парето решения состоят из трех областей.Решения в левом нижнем углу представляют собой относительно низкие метаболические затраты, но не позволяют отслеживать наблюдаемые профили крутящего момента в суставах. Производительность этих решений обычно определяется небольшими значениями F max ; минимизация этих параметров приводит к снижению метаболизма, но ослаблению мышечной силы. В правом верхнем углу представлены растворы с отличным кинетическим соответствием, но с высокими метаболическими затратами. Эти решения существенно превосходят наблюдаемые профили крутящего момента, заставляя одну или несколько мышц работать сильнее, чем это физически разумно.Эти усилия необходимы для максимального кинетического соответствия из-за недостатков данных, особенно в измерениях ЭМГ. Решения, которые представляют человека, должны находиться где-то между этими двумя крайностями, обеспечивая хорошую кинетическую пригодность при разумных метаболических затратах.
Чтобы выбрать решение в биологически приемлемой области, мы сверились с балансом метаболической энергии каждого субъекта. На рис. 5 показана полиномиальная подгонка доли метаболических затрат всего тела, потребляемых каждой смоделированной мышцей, в зависимости от кинетической подгонки (количественно выраженной средним крутящим моментом в суставе R 2 ) вдоль фронта Парето для каждого участника.Дроби представляют собой сумму по обеим сторонам и учитывают базовые расходы. Как видно из графиков, бюджет довольно постоянен для кинетической подгонки от низкого до среднего и MCOT, затем несколько мышц начинают доминировать по мере приближения к диапазону высоких R 2 и метаболических затрат. Обычно от одной до трех мышц перенапрягаются, чтобы добиться чуть лучшей кинетической подгонки, в результате чего распределение метаболических затрат значительно отличается от более разумных энергий.Физически это привело бы к утомлению рассматриваемой мышцы, или если бы она была смоделирована как намного превышающая ее фактический размер (поскольку мышечная масса принимается пропорциональной F max ). На рис. 5 показано, что группа мышц широкой мышцы бедра (ВАШ) обеспечивает наибольший вклад в метаболические затраты для каждого участника и увеличивает свою долю по мере увеличения кинетической подгонки. Это вызвано большим размером обширной мышцы живота и тенденцией оптимизатора вкладывать в нее все больше энергии, если момент разгибания колена в ранней стойке недостаточен.Учитывая важность этой мышцы для общего метаболического бюджета и ее постоянное наращивание, мы решили использовать ее для выбора оптимального решения.
Чтобы найти оптимальные решения, мы сначала аппроксимируем тенденции дробных метаболических затрат как функцию кинетической подгонки полиномом пятого порядка, как показано на рис. 5. Затем мы рассмотрели относительное изменение фракционного метаболического потребления обширной мышцы бедра Δ VAS : (17)
Здесь F VAS представляет собой долю общих метаболических затрат, приписываемых широкой мышечной ткани, как функцию среднего коэффициента детерминации кинетической подгонки ( R 2 ).Мы обнаружили, что выбор Δ VAS , opt = 0,63 в качестве порогового значения позволил нам количественно сопоставить экспериментальные метаболические затраты у четырех из пяти участников и привел к средней ошибке MCOT менее 0,01. Это пороговое значение представлено синими пунктирными вертикальными линиями на рис. 5, а черные вертикальные линии представляют погрешность экспериментального измерения. Оба набора вертикальных линий были получены путем сопоставления значений R 2 , перечисленных на оси x на фиг.5, с соответствующими метаболическими затратами в решениях, показанных на фиг.4.За выбранное оптимальное решение была принята точка с максимальной кинетической подгонкой вдоль фронта Парето, которая имела Δ VAS <Δ VAS , opt . На рис. 4 эта точка представлена в виде белого ромба на графике пространства решений каждого участника. Полные наборы параметров для этих оптимальных решений показаны в Таблице 6. Для справки список часто используемых сокращений и символов приведен в Таблице 7.
Благодарности
Мы хотим поблагодарить Питера Лоана, Брайана Амбергера и Брюса Деффенбо за их помощь в создании этой модели.Мы также хотели бы поблагодарить Дэниела Либермана за то, что он позволил нам использовать его лабораторию для сбора данных, используемых в этом исследовании.
Вклад авторов
Задумал и спроектировал эксперименты: HH JM. Выполнял эксперименты: JM. Проанализированы данные: JM. Предоставленные реагенты / материалы / инструменты анализа: JM HH. Написал статью: JM HH.
Ссылки
- 1. Chao EY, An KN. Графическая интерпретация решения избыточной проблемы биомеханики.J Biomech. Engng 1978; 100 (3): 159–167.
- 2. Ямагути Г., Морана Д., Дж. С. Вычислительно эффективный метод решения избыточной проблемы в биомеханике. J Biomech. 1995; 28: 999–1005. pmid: 7673268
- 3. Кандел Э. Р., Шварц Дж. Х., Джесселл ТМ. Принципы неврологии. 4-е изд. Нью-Йорк, штат Нью-Йорк: Макгроу-Хилл; 2000.
- 4. Клэнси Э.А., Морин Э.Л., Мерлетти Р. Проблемы отбора проб, шумоподавления и оценки амплитуды в поверхностной электромиографии.Журнал электромиографии и кинезиологии. 2002; 12 (1): 1–16. pmid: 11804807
- 5. Хоган Н., Манн Р. В.. Обработка миоэлектрического сигнала: оптимальная оценка применительно к электромиографии — Часть II: экспериментальная демонстрация оптимальной производительности миопроцессора. IEEE Transactions по биомедицинской инженерии. 1980. 27 (7): 396–410. pmid: 7409805
- 6. Хоган Н., Манн Р.У. Обработка миоэлектрических сигналов: оптимальная оценка применительно к электромиографии — Часть I: Создание оптимального миопроцессора.IEEE Transactions по биомедицинской инженерии. 1980. 27 (7): 382–395. pmid: 7409804
- 7. Фукунага Т., Кубо К., Каваками Ю., Фукасиро С., Канехиса Н., Маганарис С. Н.. Поведение мышечного сухожилия человека при ходьбе in vivo. Труды Биологических наук / Королевское общество. 2001 февраль; 268 (1464): 229–33. Доступно по адресу: https://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1088596&tool=pmcentrez&rendertype=abstract. pmid: 11217891
- 8. Исикава М., Коми П.В., Грей М.Дж., Лепола В., Брюггеманн Г.П.Взаимодействие мышц и сухожилий и использование упругой энергии при ходьбе человека. Журнал прикладной физиологии (Bethesda, Мэриленд: 1985). 2005 август; 99 (2): 603–8. Доступно по адресу: https://www.ncbi.nlm.nih.gov/pubmed/15845776.
- 9. Lichtwark GA, Bougoulias K, Wilson AM. Длина мышечного пучка и ряда упругих элементов изменяется по длине икроножной мышцы человека при ходьбе и беге. Журнал биомеханики. 2007. 40: 157–164. pmid: 16364330
- 10. Chleboun GS.Изменение длины пучка передней и латеральной большеберцовой мышцы человека во время ходьбы. Журнал ортопедии и спортивной физиотерапии. Июль 2007 г.; 37 (7): 372–379. Доступно по адресу: https://www.jospt.org/issues/id.1286/article_detail.asp. pmid: 17710906
- 11. Рубенсон Дж., Пирес Нью-Джерси, Лой Х.О., Пиннигер Дж. Дж., Шеннон Д. Г.. На подъеме: рабочая длина камбаловидной мышцы сохраняется на восходящем участке кривой «сила-длина» в механике походки человека. Журнал экспериментальной биологии 2012; 215: 3539–3551.pmid: 22771749
- 12. Фаррис Д.Ф., Савицкий Г.С. Поведение силы-скорости медиальной икроножной мышцы человека изменяется в зависимости от скорости передвижения и походки. Proc Natl Acad Sci U S A. 2012 Янв; 109 (3): 977–82. pmid: 22219360
- 13. Лай А., Лихтварк Г.А., Шахе А.Г., Линь Ю.С., Браун НАТ, Панди М.Г. In vivo поведение камбаловидной мышцы человека при увеличении скорости ходьбы и бега. Журнал прикладной физиологии. 2015 Май; 118 (10): 1266–1275. pmid: 25814636
- 14.Delp SL, займ JP. Графическая система программного обеспечения для разработки и анализа моделей костно-мышечных структур. Компьютеры в биологии и медицине. 1995. 25 (1): 21–34. pmid: 7600758
- 15. Zajac FE. Мышцы и сухожилия: свойства, модели, масштабирование и применение в биомеханике и моторном управлении. Критические обзоры в биомедицинской инженерии. 1989. 17 (4): 359–411. pmid: 2676342
- 16. Винтерс Дж. М.. Модели мышц на основе холма: перспектива системной инженерии.В: Winters JM, Woo SLY, редакторы. Множественные мышечные системы: биомеханика и организация движений. Springer-Verlag; 1990. стр. 69–93.
- 17. Андерсон ФК, Панди МГ. Динамическая оптимизация ходьбы человека. Журнал биомеханической инженерии. 2001; 123 (5): 381. Доступно по ссылке: https://link.aip.org/link/JBENDY/v123/i5/p381/s1&Agg=doi. pmid: 11601721
- 18. Нептун Р. Р., Сасаки К., Каутц Са. Влияние скорости ходьбы на функцию мышц и механическую энергетику.Походка и поза. Июль 2008 г .; 28 (1): 135–43. Доступно по адресу: https://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2409271&tool=pmcentrez&rendertype=abstract.
- 19. Гейер Х., Герр Х. Модель мышечного рефлекса, которая кодирует принципы механики ног, дает человеческую динамику ходьбы и мышечную активность. Операции IEEE по нейронным системам и реабилитационной инженерии: публикация Общества инженеров IEEE в медицине и биологии. 2010 июн; 18 (3): 263–73.Доступно по адресу: https://www.ncbi.nlm.nih.gov/pubmed/20378480. Код доступен по адресу: https://www.cs.cmu.edu/~hgeyer/Software/Neuromuscular%20Model/Geyer%20Neuromuscular%20Model.zip.
- 20. Lichtwark G, Wilson A. Оптимальная длина мышечных пучков и жесткость сухожилий для максимальной эффективности икроножной мышцы во время ходьбы и бега человека. J Theor Biol. 2008. 252 (4): 662–73. pmid: 18374362
- 21. Хилл А.В. Теплота сокращения и динамические константы мышц.Труды Лондонского королевского общества, серия B, биологические науки. 1938; 126 (843): 136–195.
- 22. Робертс Т. Комплексная функция мышц и сухожилий при движении. Comp Biochem Physiol A Mol Integr Physiol. 2002; 133: 1087–1099. pmid: 12485693
- 23. Кришнасвами П., Браун Э. Н., Герр Х. М.. Модель ноги человека предсказывает морфологию, состояние, роль и энергетику голеностопных мышц и сухожилий при ходьбе. Вычислительная биология PLoS. 2011 Март; 7 (3): e1001107. Доступно по адресу: https: // www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3060164&tool=pmcentrez&rendertype=abstract. pmid: 21445231
- 24. Умбергер Б.Р., Герритсен КГМ, Мартин П.Е. Модель расхода энергии в мышцах человека. Компьютерные методы в биомеханике и биомедицинской инженерии. 2003 Апрель; 6 (2): 99–111. Доступно по адресу: https://www.ncbi.nlm.nih.gov/pubmed/12745424. pmid: 12745424
- 25. Sanger TD. Байесовская фильтрация миоэлектрических сигналов. Журнал нейрофизиологии. 2007 февраль; 97 (2): 1839–45.Доступно по адресу: https://www.ncbi.nlm.nih.gov/pubmed/17182908. Код доступен по адресу: https://www.kidsmove.org/bayesemgdemo.html pmid: 17182908
- 26. He J, Levine WS, Loeb GE. Усиление обратной связи для коррекции небольших возмущений позы стоя. IEEE Trans. по автоматическому управлению. 1991; 36: 322–332.
- 27. Рааш CC, Заяц Ф. Е., Ма Б., Левин В. С.. Мышечная координация при педалировании на максимальной скорости. Журнал биомеханики. 1997. 30: 595–602. pmid:
- 93
- 28.Перри Дж. Анализ походки. 1-е изд. Thorofare, NJ 08086-9447: SLACK Incorporated; 1992.
- 29. Эндо К., Герр Х. Модель функции сухожилий и мышц при ходьбе человека с самостоятельно выбранной скоростью. Операции IEEE по нейронным системам и реабилитационной инженерии: публикация Общества инженеров IEEE в медицине и биологии. 2013; 22: 352–62.
- 30. Деб К. Многоцелевая оптимизация с использованием эволюционных алгоритмов. Хобокен, штат Нью-Джерси, 07030-5774: John Wiley & Sons; 2001 г.
- 31. Сартори М., Реджиани М., Фарина Д., Ллойд Д.Г. ЭМГ-управляемая прямая динамическая оценка мышечной силы и суставного момента о нескольких степенях свободы в нижней конечности человека. PLoS ONE. 2012; 7 (12): e52618. pmid: 23300725
- 32. McMahon TA. Мышцы, рефлексы и движение. Принстон, Нью-Джерси: Издательство Принстонского университета; 1984.
- 33. Савицкий Г., Феррис Д. Механика и энергетика горизонтальной ходьбы с экзоскелетами голеностопного сустава с электроприводом.J Exp Biol. 2008; 211: 1402–1413. pmid: 18424674
- 34. Umberger BR. Стоимость фазы стойки и поворота при ходьбе человека. Журнал Королевского общества, Интерфейс / Королевское общество. 2010 Сен; 7 (50): 1329–40. Доступно по адресу: https://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2894890&tool=pmcentrez&rendertype=abstract. pmid: 20356877
- 35. Арнольд Э.М., Дельп С.Л. Рабочие длины волокон мышц нижних конечностей человека при ходьбе. Фил. Пер. R. Soc.В 2011; 366: 1530–1539. pmid: 21502124
- 36. Арнольд Э.М., Хамнер С.Р., Сет А., Миллард М., Delp SL. Как длина и скорость мышечных волокон влияют на выработку мышечной силы при ходьбе и беге человека с разной скоростью. Журнал экспериментальной биологии, 2013; 216: 2150–2160. pmid: 23470656
- 37. Бьюкенен Т.С., Ллойд Д.Г., Манал К., Безье Т.Ф. Оценка мышечных сил и моментов в суставах с использованием модели прямой-обратной динамики. Med. Sci. Спортивные упражнения. 2005; 37: 1911–1916.
- 38. Ллойд Д.Г., Безье Т.Ф. Модель опорно-двигательного аппарата на основе ЭМГ для оценки мышечных сил и моментов в коленных суставах in vivo. J. Biomech. 2003; 36: 765–776. pmid: 12742444
- 39. Коркос Д.М., Готтлиб Г.Л., Латаш М.Л., Алмейда Г.Л., Агарвал Г.К. Электромеханическая задержка: экспериментальный артефакт. J. Electromyogr. Кинезиол. 1992; 2: 59–68. pmid: 20719599
- 40. Гимарайнш А.С., Херцог В., Аллингер Т.Л., Чжан Ю.Т. Взаимосвязь ЭМГ-силы камбаловидной мышцы кошки и ее связь с условиями сокращения во время передвижения.J. Exp. Биол. 1995; 198: 975–987. pmid: 7730759
- 41. Дэви Д.Т., Оду М.Л. Метод динамической оптимизации для прогнозирования мышечных сил в фазе качания походки. J. Biomech. 1987; 20: 187–201. pmid: 3571299
- 42. О’Рахилли Р., Мюллер Ф., Карпентер С., Свенсон Р. Основы анатомии человека Авторские права O’Rahilly 2004 г. Доступно по адресу: https://www.dartmouth.edu/~humananatomy/.
- 43. Пандольф К.Б., Гивони Б., Гольдман РФ. Прогнозирование расхода энергии с нагрузками при очень медленной стойке или ходьбе.Научно-исследовательский институт экологической медицины армии США. Отчет № М 3/77, 1976.
- 44. Эндо К., Палуска Д., Герр Х. Квазипассивная модель функции ног человека при ходьбе по ровной поверхности. 2006 Международная конференция IEEE / RSJ по интеллектуальным роботам и системам. 2006 Oct; п. 4935–4939. Доступно по адресу: https://ieeexplore.ieee.org/lpdocs/epic03/wrapper.htm?arnumber=4059202.
- 45. Хилл А.В. Эффективность развития механической силы при сокращении мышц и ее связь с нагрузкой.Труды Лондонского королевского общества, серия B, биологические науки. 1964; 159 (975).
- 46. Сонг С., Десаи Р., Гейер Х. Интеграция адаптивного управления махом в нейромышечную модель ходьбы человека. Материалы конференции: Ежегодная международная конференция общества инженеров в медицине и биологии IEEE, Конференция в Осаке, Япония. 2013 июл; с. 4915–4918.
- 47. Сонг, С., Гейер, Х. Регулирование скорости и создание больших переходов скорости в нейромышечной модели ходьбы человека.2012 Международная конференция IEEE по робототехнике и автоматизации. 2012 Май; с. 511–516. Доступно по адресу: https://ieeexplore.ieee.org/lpdocs/epic03/wrapper.htm?arnumber=6225307.
- 48. Ван Дж. М., Хамнер С. Р., Делп С. Л., Колтун В. Оптимизация контроллеров движения с использованием биологических приводов и объективов. Транзакции ACM на графике (Proc SIGGRAPH 2012). 2012; 31 (4).
- 49. Сонг С., Гейер Х. Обобщение модели мышечно-рефлексного контроля на трехмерную ходьбу. Материалы конференции: Ежегодная международная конференция общества инженеров в медицине и биологии IEEE, Конференция в Осаке, Япония.2013 июл; с. 7463–7466.
- 50. Brockway JM. Вывод формул, используемых для расчета расхода энергии у человека. Питание человека Клиническое питание. 1987 ноя; 41 (6): 463–71. Доступно по адресу: https://www.ncbi.nlm.nih.gov/pubmed/3429265. pmid: 3429265
- 51. Крам Дж. Р., Касман Г. С., Хольц Дж. Введение в поверхностную электромиографию 1-е изд. Гейтерсбург, Мэриленд 20878: Аспен Паблишерс; 1998.
- 52. Хатце Х. Модель миокибернетического контроля скелетных мышц.Биологическая кибернетика. 1977; 25: 103–119. pmid: 836914
стен из древних костей человеческих ног | Умные новости
Археологи обнаружили девять стен из человеческих костей под собором Святого Бавона в Генте, Бельгия. © Авторское право Ruben Willaert bvbaБолее 100 миллиардов человек умерли с тех пор, как линия Homo sapiens впервые появилась примерно 50 000 лет назад. И с таким количеством тел, которые нужно хоронить, цивилизациям постоянно не хватало удобных мест для их захоронения.
Но люди изобретательны. Чтобы освободить место для новых могил, жители Гента, Бельгия, по-видимому, выкопали несколько старых могил и собрали кости, погребенные внутри, в девять стен. Как сообщает Алан Хоуп для газеты Brussels Times , археологи, проводящие раскопки перед строительством нового центра для посетителей собора, теперь обнаружили остатки этих ужасных архитектурных подвигов.
Обнаруженные под собором Святого Баво, где находится знаменитый Гентский алтарь , недавно обнаруженные костяные стены, вероятно, служили той же цели, что и катакомбы Парижа, предоставляя второй дом для останков скелетов, вытесненных из старых кладбищ.
Так называемые оссуарии не редкость, но обычно эти камеры проявляются в виде рыхлых ямок, заполненных костями. «Тщательно построенные стены представляют собой первые в Бельгии», — пишет Яник де Гриз, археолог, руководивший раскопками, в электронном письме к Лауре Геггель из Live Science .
Де Гриз добавляет: «Мы никогда не видели структур, таких как стены, которые намеренно построены из человеческих костей».
Состоящие в основном из костей бедра и голени взрослого человека, стены также перемежаются частично раздробленными черепами.Хотя полностью неповрежденные скелеты предположительно служили основным кормом строителей, самые маленькие и хрупкие кости, такие как ребра, позвонки и все, что когда-то принадлежало маленькому ребенку, не вошли в конечный продукт — возможно, потому, что строители не смогли » не беспокойтесь о том, чтобы их собрать. Также заметно отсутствуют кости рук, которые, при прочих равных, должны были быть достаточно прочными, чтобы их можно было использовать в конструкциях.
По неизвестным причинам эти конкретные останки оказались неподходящими, возможно, потому, что они физически не соответствовали размеру костей ног или потому, что они не имели правильного духовного значения, как говорит де Гриз Live Science .
В слое грязи над стенами находятся дополнительные человеческие останки, сохранившиеся в виде полных скелетов, намекая на то, что более недавнее кладбище — то, которое, возможно, бесцеремонно загрузило жителей своего предшественника, — оставалось в использовании в течение некоторого времени после того, как были построены костлявые баррикады. , согласно Brussels Times .
Согласно Live Science , радиоуглеродное датирование предполагает, что кости в стенах принадлежали людям, умершим во второй половине 15 века.Сами стены, вероятно, относятся к 1600-м или началу 1700-х годов.
Хотя скелетные структуры могут показаться современным читателям бессердечными, де Гриз сообщает Brussels Times , что «верующие верили в воскресение тела. Кости считались самой важной частью ».
Учитывая, что альтернативой могло быть просто выбросить останки, возведение стен было, возможно, не столько неуважительным показом, сколько чрезвычайно эффективной данью уважения давно ушедшим.
В то время как другие оссуарии в Европе приобрели известность как туристические достопримечательности, костяные стены Гента кажутся обреченными на загробную жизнь вдали от всеобщего внимания. Как сказал де Гриз в интервью Live Science , останки будут удалены по мере продолжения строительства и переведены в исследовательские центры Гентского университета, которые в конечном итоге могут стать их последним пристанищем.
Археология Кости Крутые находки Смерть Европейская историяРекомендованные видео
Аппендикулярные мышцы тазового пояса и нижних конечностей — анатомия и физиология
Цели обучения
К концу этого раздела вы сможете:
- Определите аппендикулярные мышцы тазового пояса и нижней конечности
- Определить движение и функцию тазового пояса и нижней конечности
Аппендикулярные мышцы нижней части тела и стабилизируют тазовый пояс, который служит основой для нижних конечностей.Для сравнения, в грудном поясе движения гораздо больше, чем в тазовом. Движение тазового пояса очень мало из-за его соединения с крестцом в основании осевого скелета. Тазовый пояс имеет меньший диапазон движений, потому что он был разработан для стабилизации и поддержки тела.
Мышцы бедра
Что бы произошло, если бы тазовый пояс, соединяющий нижние конечности с туловищем, имел такой же диапазон движений, что и грудной пояс? Во-первых, ходьба потребовала бы больше энергии, если бы головки бедер не были закреплены в вертлужной впадине таза.Центр тяжести тела находится в области таза. Если бы центр тяжести не оставался неизменным, встать было бы также трудно. Таким образом, то, что мышцам ног не хватает диапазона движений и гибкости, они компенсируют размером и мощностью, облегчая стабилизацию, осанку и движения тела.
Ягодичные мышцы, двигающие бедро
Большинство мышц, которые входят в бедренную кость (бедренную кость) и перемещают ее, берут начало в тазовом поясе. Большая поясничная мышца и подвздошная мышца составляют группу подвздошно-поясничной мышцы.Одними из самых крупных и мощных мышц тела являются ягодичные мышцы или ягодичная группа. Большая ягодичная мышца самая большая; Глубоко от большой ягодичной мышцы находится средняя ягодичная мышца, а глубоко от средней ягодичной мышцы — минимальная ягодичная мышца, самая маленькая из трех ((Рисунок) и (Рисунок)).
Мышцы бедра и бедра
Большие и мощные мышцы бедра, которые двигают бедренную кость, обычно берут начало в тазовом поясе и переходят в бедренную кость. Мышцы, которые двигают голень, обычно берут начало на бедре и вставляются в кости коленного сустава.Передние мышцы бедра расширяют голень, но также помогают сгибать бедро. Задние мышцы бедра сгибают голень, но также помогают разгибать бедро. Комбинация ягодичных мышц и мышц бедра также приводит, отводит и вращает бедро и голень.
Ягодичные мышцы, двигающие бедро
Tenor fascia latae — толстая квадратная мышца в верхней части боковой поверхности бедра. Он действует как синергист средней ягодичной мышцы и подвздошно-поясничной мышцы при сгибании и отведении бедра.Это также помогает стабилизировать латеральную сторону колена, натягивая подвздошно-большеберцовый тракт (бандаж), делая его туго натянутым. Глубоко от большой ягодичной мышцы грушевидная мышца, внутренняя запирательная мышца, наружная запирательная мышца, верхний гемеллус, нижний гемеллюс и квадратная мышца бедра вращают бедренную кость в боковом направлении.
Длинная приводящая мышца, короткая приводящая мышца и большая приводящая мышца могут поворачивать бедро как медиально, так и латерально, в зависимости от положения стопы. Длинная приводящая мышца сгибает бедро, а большая приводящая мышца разгибает его.Грудная мышца приводит и сгибает бедренную кость в тазобедренном суставе. Грудная мышца расположена в бедренном треугольнике, который образуется на стыке бедра и голени, а также включает бедренный нерв, бедренную артерию, бедренную вену и глубокие паховые лимфатические узлы.
Мышцы бедра, двигающие бедренную, большеберцовую и малоберцовую кости
Глубокая фасция бедра разделяет его на медиальный, передний и задний отделы (см. (Рисунок) и (Рисунок)). Мышцы медиального отдела бедра отвечают за приведение бедра к бедру.Наряду с длинной приводящей мышцей, короткой приводящей мышцей, большой приводящей мышцей и грудной клеткой, тонкая мышца в виде ремня аддуктирует бедро в дополнение к сгибанию ноги в колене.
Мышцы бедра, двигающие бедренную, большеберцовую и малоберцовую кости
Мышцы переднего отдела бедра сгибают бедро и разгибают ногу. Этот отсек содержит группу четырехглавой мышцы бедра, которая на самом деле состоит из четырех мышц, которые разгибают и стабилизируют колено. Прямая мышца бедра находится на передней поверхности бедра, латеральная широкая мышца бедра находится на боковой поверхности бедра, медиальная широкая мышца бедра находится на медиальной стороне бедра, а средняя широкая мышца бедра находится между латеральной широкой мышцей бедра и медиальной широкой мышцей бедра и глубокой. к прямой мышце бедра.Общее для всех четырех сухожилий — это сухожилие четырехглавой мышцы (сухожилие надколенника), которое входит в надколенник и продолжается под ним как связка надколенника. Связка надколенника прикрепляется к бугристости большеберцовой кости. Помимо четырехглавой мышцы бедра, портняжная мышца представляет собой ленточноподобную мышцу, которая простирается от передней верхней подвздошной ости до медиальной стороны проксимального отдела большеберцовой кости. Эта универсальная мышца сгибает ногу в колене и сгибает, отводит и поворачивает ногу в сторону бедра. Эта мышца позволяет нам сидеть, скрестив ноги.
Задний отдел бедра включает мышцы, которые сгибают ногу и разгибают бедро. Три длинные мышцы задней части колена — это группа подколенного сухожилия, которая сгибает колено. Это двуглавая мышца бедра, полусухожильная и полуперепончатая мышца. Сухожилия этих мышц образуют подколенную ямку, ромбовидное пространство на задней стороне колена.
Мышцы, двигающие стопы и пальцы ног
Подобно мышцам бедра, мышцы ноги разделены глубокой фасцией на отсеки, хотя в ноге их три: передняя, боковая и задняя ((Рисунок) и (Рисунок)).
Мышцы голени
Мышцы переднего отдела голени обычно отвечают за тыльное сгибание, а мышцы заднего отдела голени обычно отвечают за подошвенное сгибание. Боковые и медиальные мышцы в обоих отделах инвертируют, разворачивают и вращают стопу.
Мышцы, двигающие стопы и пальцы ног
Мышцы в переднем отделе голени: передняя большеберцовая мышца, длинная и толстая мышца на боковой поверхности большеберцовой кости, длинный разгибатель большого пальца стопы глубоко под ней и длинный разгибатель пальцев сбоку от нее — все они способствуют поднимать переднюю часть стопы при сокращении.Тертиус малоберцовой кости, небольшая мышца, которая берет начало на передней поверхности малоберцовой кости, связана с длинным разгибателем пальцев и иногда сливается с ним, но присутствует не у всех людей. Толстые связки соединительной ткани, называемые верхним удерживателем разгибателей (поперечная связка голеностопного сустава) и нижним удерживателем разгибателей, удерживают сухожилия этих мышц на месте во время тыльного сгибания.
Боковой отсек голени включает две мышцы: длинную малоберцовую мышцу (peroneus longus) и малоберцовую мышцу (малоберцовую мышцу).Все поверхностные мышцы в заднем отделе ноги вставляются в пяточное сухожилие (ахиллово сухожилие), сильное сухожилие, которое входит в пяточную кость голеностопного сустава. Мышцы в этом отсеке большие и сильные, они удерживают человека в вертикальном положении. Самая поверхностная и видимая мышца голени — икроножная мышца. Глубоко от икроножной мышцы проходит широкая плоская камбаловидная мышца. Плантарис проходит между ними наискосок; у некоторых людей может быть две из этих мышц, в то время как подошва не наблюдается примерно в семи процентах других случаев вскрытия трупа.Сухожилие подошвенной мышцы является желательной заменой широкой фасции при герниопластике, трансплантации сухожилий и восстановлении связок. В заднем отделе голени также есть четыре глубоких мышцы: подколенная мышца, длинный сгибатель пальцев, длинный сгибатель большого пальца стопы и задняя большеберцовая мышца.
Стопа также имеет собственные мышцы, которые берут начало и вставляются в нее (аналогично внутренним мышцам руки). Эти мышцы в первую очередь обеспечивают поддержку стопы и ее свода, а также способствуют движениям пальцев ног ((Рисунок) и (Рисунок)).Основной опорой продольного свода стопы является глубокая фасция, называемая подошвенным апоневрозом, которая проходит от пяточной кости до пальцев стопы (воспаление этой ткани является причиной «подошвенного фасциита», который может поражать бегунов. Стопа состоит из двух групп. Дорсальная группа включает только одну мышцу, brevis Extensor digitorum. Вторая группа — подошвенная группа, которая состоит из четырех слоев, начиная с самого поверхностного.
Внутренние мышцы стопы
Мышцы тыльной стороны стопы (а) обычно разгибают пальцы ног, в то время как мышцы подошвенной стороны стопы (b, c, d) обычно сгибают пальцы ног.Подошвенные мышцы состоят из трех слоев, обеспечивая стопе силу, уравновешивающую вес тела. На этой диаграмме эти три слоя показаны с точки зрения подошвы, начиная с самого нижнего слоя непосредственно под подошвенной кожей стопы (b) и заканчивая самым верхним слоем (d), расположенным чуть ниже стопы и пальца ноги. кости.
Внутренние мышцы стопы
Обзор главы
Тазовый пояс прикрепляет ноги к осевому каркасу.В тазобедренном суставе тазовый пояс соединяется с ногой. Бедро соединено с тазовым поясом множеством мышц. В ягодичной области большая поясничная мышца и подвздошная мышца образуют подвздошно-поясничную мышцу. Большая и сильная ягодичная мышца большая, средняя и минимальная ягодичные мышцы расширяют и отводят бедро. Наряду с большой ягодичной мышцей, напрягающая широкая фасция мышцы образует подвздошно-большеберцовый тракт. Латеральными вращающими элементами бедренной кости в области бедра являются грушевидная мышца, внутренняя запирательная мышца, внешняя запирательная мышца, верхний гемеллус, нижний гемеллюс и квадратная мышца бедра.На медиальной части бедра длинная приводящая мышца, короткая приводящая мышца и большая приводящая мышца приводят бедро и вращают его кнутри. Грудная мышца приводит и сгибает бедро в области бедра.
Мышцы бедра, двигающие бедренную, большеберцовую и малоберцовую кости, делятся на медиальный, передний и задний отделы. Медиальный отсек включает аддукторы, грудную мышцу и тонкую мышцу. Передний отдел включает четырехглавую мышцу бедра, сухожилие четырехглавой мышцы, связку надколенника и портняжную мышцу.Четырехглавая мышца бедра состоит из четырех мышц: прямой мышцы бедра, латеральной широкой мышцы бедра, средней широкой мышцы бедра и промежуточной широкой мышцы бедра, которые вместе расширяют колено. Задний отдел бедра включает подколенные сухожилия: двуглавая мышца бедра, полусухожильная и полуперепончатая мышцы, которые сгибают колено.
Мышцы ноги, двигающие стопы и пальцы ног, делятся на передний, боковой, поверхностный и глубокий задний отделы. Передний отдел включает переднюю большеберцовую мышцу, длинный разгибатель большого пальца стопы, длинный разгибатель пальцев и третичную малоберцовую мышцу.В боковом отделе находятся длинная малоберцовая мышца (малоберцовая мышца) и короткая малоберцовая мышца (малоберцовая мышца). Поверхностный задний отдел включает икроножную, камбаловидную и подошвенную мышцу; а в глубоком заднем отделе находятся подколенная мышца, задняя большеберцовая мышца, длинный сгибатель пальцев и длинный сгибатель большого пальца стопы.
Обзорные вопросы
Большая группа мышц, которая прикрепляет ногу к тазовому поясу и производит разгибание тазобедренного сустава, — это группа ________.
- ягодичная
- обтуратор
- приводящая мышца
- похититель
Какая мышца производит движение, позволяющее скрестить ноги?
- большая ягодичная мышца
- грушевидная
- грацилис
- портняжник
Какая самая большая мышца голени?
- камбала
- икроножная мышца
- передняя большеберцовая мышца
- Задняя большеберцовая мышца
До какой из следующих мышц глубина промежуточной широкой мышцы бедра?
- двуглавая мышца бедра
- прямая мышца бедра
- Вастус медиальная
- Вастус латеральная
Вопросы о критическом мышлении
Какие мышцы образуют подколенные сухожилия? Как они работают вместе?
Двуглавая, полуперепончатая и полусухожильная мышцы бедра образуют подколенные сухожилия.Подколенные сухожилия сгибают ногу в коленном суставе.
Какие мышцы образуют четырехглавую мышцу? Как они работают вместе?
Прямая мышца бедра, медиальная широкая мышца бедра, латеральная широкая мышца бедра и промежуточная широкая мышца бедра образуют четырехглавую мышцу. Четырехглавая мышца разгибает ногу в коленном суставе.
Глоссарий
- приводящая мышца короткая
- мышца, приводящая бедро и вращающая его кнутри
- длинная приводящая мышца
- мышца, приводящая, вращающая кнутри и сгибающая бедро
- большая приводящая мышца
- мышца с передним пучком, который аддуктирует, медиально вращает и сгибает бедро, и задним пучком, который способствует разгибанию бедра
- передний отдел ноги
- область, включающая мышцы, сгибающие стопу сзади
- передний отдел бедра
- область, включающая мышцы, сгибающие бедро и разгибающие ногу
- двуглавая мышца бедра
- мышца подколенного сухожилия
- пяточное сухожилие
- (также ахиллово сухожилие) сильное сухожилие, которое вставляется в пяточную кость голеностопного сустава
- спинная группа
- область, которая включает короткий разгибатель пальцев
- короткий разгибатель пальцев
- мышца, разгибающая пальцы ног
- длинный разгибатель пальцев
- мышца латеральнее передней большеберцовой мышцы
- длинный разгибатель большого пальца стопы
- мышца, которая частично проходит глубоко от передней большеберцовой мышцы и длинного разгибателя пальцев
- бедренный треугольник
- Область, образованная на стыке бедра и голени, включает грудную мышцу, бедренный нерв, бедренную артерию, бедренную вену и глубокие паховые лимфатические узлы
- fibularis brevis
- (также, малоберцовая мышца) подошвенная мышца, сгибающая стопу в голеностопном суставе и выворачивающая ее в межплюсневых суставах
- длинная малоберцовая мышца
- (также длинная малоберцовая мышца) мышца, которая сгибает стопу в голеностопном суставе и выворачивает ее в межплюсневых суставах
- третичная малоберцовая мышца
- маленькая мышца, связанная с длинным разгибателем пальцев
- длинный сгибатель пальцев
- мышца, которая сгибает четыре мизинца
- длинный сгибатель большого пальца стопы
- мышца, сгибающая большой палец ноги
- икроножная мышца
- Самая поверхностная мышца голени
- ягодичная группа
- Группа мышц, которая разгибает, сгибает, вращает, аддукт и отводит бедро
- большая ягодичная мышца
- самая большая из ягодичных мышц, расширяющая бедро
- средняя ягодичная мышца
- мышца глубоко от большой ягодичной мышцы, которая отводит бедро у бедра
- малая ягодичная мышца
- наименьшая из ягодичных мышц и глубокая до средней ягодичной мышцы
- грацилис
- мышца, приводящая бедро и сгибающая ногу в колене
- группа подколенного сухожилия
- три длинные мышцы задней части голени
- подвздошная кость
- мышца, которая вместе с большой поясничной мышцей составляет подвздошно-поясничную мышцу
- подвздошно-поясничная группа
- Группа мышц, состоящая из подвздошной мышцы и большой поясничной мышцы, которая сгибает бедро в бедре, вращает его в стороны и сгибает туловище на бедре
- подвздошно-большеберцовый тракт
- мышца, прикрепляющаяся к большеберцовой кости; состоит из большой ягодичной мышцы и соединительной ткани растяжения широкой фасции
- удерживатель нижнего разгибателя
- крестообразная связка голеностопного сустава
- inferior gemellus
- мышца глубоко от большой ягодичной мышцы на боковой поверхности бедра, которая вращает бедро в боковом направлении
- боковой отсек ноги
- область, которая включает длинную малоберцовую мышцу (малоберцовую мышцу) и короткую малоберцовую мышцу (малоберцовую мышцу) и связанные с ними кровеносные сосуды и нервы
- медиальный отдел бедра
- область, которая включает длинную приводящую мышцу, короткую приводящую мышцу, большую приводящую мышцу, грудную мышцу, тонкую мышцу и связанные с ними кровеносные сосуды и нервы
- наружная запирательная часть
- мышца глубоко от большой ягодичной мышцы на боковой поверхности бедра, которая вращает бедро в боковом направлении
- внутренняя запирательная мышца
- мышца глубоко от большой ягодичной мышцы на боковой поверхности бедра, которая вращает бедро в боковом направлении
- связка надколенника
- разгибание сухожилия четырехглавой мышцы ниже надколенника
- грудная клетка
- мышца, отводящая и сгибающая бедро в области бедра
- тазовый пояс
- бедра, опора для нижней конечности
- грушевидная
- мышца глубоко от большой ягодичной мышцы на боковой поверхности бедра, которая вращает бедро в боковом направлении
- подошвенный апоневроз
- мышца, поддерживающая продольный свод стопы
- подошвенная группа
- четырехслойная группа собственных мышц стопы
- плантарис
- мышца, которая проходит наискось между икроножной и камбаловидной мышцами
- подколенная ямка
- ромбовидное пространство на тыльной стороне колена
- подколенная
- мышца, которая сгибает ногу в колене и образует дно подколенной ямки
- задний отдел голени
- область, которая включает поверхностную икроножную, камбаловидную и подошвенную мышцу, а также глубокую подколенную мышцу, длинный сгибатель пальцев, длинный сгибатель большого пальца стопы и заднюю большеберцовую мышцу
- задний отдел бедра
- область, включающая мышцы, сгибающие ногу и разгибающие бедро
- поясничная большая
- мышца, которая вместе с подвздошной костью составляет подвздошно-поясничную мышцу
- квадратная мышца бедра
- мышца глубоко от большой ягодичной мышцы на боковой поверхности бедра, которая вращает бедро в боковом направлении
- группа четырехглавой мышцы бедра
- четыре мышцы, которые разгибают и стабилизируют колено
- Сухожилие четырехглавой мышцы
- (также сухожилие надколенника) сухожилие, общее для всех четырех мышц четырехглавой мышцы, вставляется в надколенник
- прямая мышца бедра
- Четырехглавая мышца передней поверхности бедра
- портной
- ленточноподобная мышца, которая сгибает, отводит и поворачивает ногу в сторону бедра
- полуперепончатая кость
- мышца подколенного сухожилия
- полусухожильная
- мышца подколенного сухожилия
- камбаловидная
- широкая, плоская мышца глубоко до икроножной мышцы
- удерживатель верхнего разгибателя
- поперечная связка голеностопного сустава
- Superior gemellus
- мышца глубоко от большой ягодичной мышцы на боковой поверхности бедра, которая вращает бедро в боковом направлении
- тензор широкой фасции
- мышца, сгибающая и отводящая бедро
- передняя большеберцовая мышца
- мышца, расположенная на боковой поверхности большеберцовой кости
- задняя большеберцовая мышца
- мышца, которая подошвенно сгибает и переворачивает стопу
- вастус промежуточный
- Четырехглавая мышца, которая находится между латеральной широкой мышцей бедра и медиальной широкой мышцей бедра и проходит глубоко в прямую мышцу бедра
- вастус латеральная
- Четырехглавая мышца на боковой поверхности бедра
- обширная медиальная мышца
- Четырехглавая мышца на медиальной стороне бедра
Анатомия ноги — значение, части и часто задаваемые вопросы
Анатомия ноги означает исследование ноги человеческого тела.Две нижние конечности тела обеспечивают поддержку и широкий диапазон движений.
Нога состоит из пяти областей, они следующие
-
Верхняя нога
-
Колено
-
Голень
-
Голеностопный сустав
-
Мышцы стопы
4 Нога
ряда длинных мышц, которые помогают растягивать дистанции. Ноги человека — это нижние конечности тела, которые обеспечивают движение и поддержку человека.Строение мышц ног зависит от части ноги. Давайте узнаем о других типах мышц ног, основанных на таких частях ног, как колено, лодыжка, ступни и бедра.Части мышц ног
Анатомию ног можно разделить на мышцы верхней и нижней части ног.
1. Мышцы верхней части ног
Верхняя часть ног состоит из трех основных мышц:
подколенных сухожилий, четырехглавой мышцы и приводящих мышц.
A. Подколенное сухожилие: подколенные сухожилия — это те три мышцы, которые расположены в задней части бедер.Эти мышцы помогают нам сгибать и сгибать колени. Эти три мышцы подколенного сухожилия — это полуперепончатая, полусухожильная и двуглавая мышцы бедра.
Полуперепончатая мышца соединяется с тазом и большеберцовой костью посредством сухожилий. Эти мышцы помогают сгибаться и вращаться. Эти мышцы помогают вращать голень на бедре, когда колено сгибается, и вращать бедро, когда бедра растянуты. Мышцы двуглавой мышцы бедра помогают растягивать бедро, сгибать колено и вращать колени вне тела.
B. Четырехглавая мышца: четырехглавая мышца — это четыре мышцы, расположенные в передней части бедер. Четырехглавая мышца помогает разгибать колено из-за степени сгибания. Четыре четырехглавой мышцы — это латеральная широкая мышца бедра, медиальная широкая мышца бедра и прямая мышца бедра.
Боковая широкая мышца бедра укрепляет голень и помогает нам подняться на корточки. Что касается коленных мышц, то медиальная широкая мышца бедра и промежуточная широкая мышца бедра помогают нам разгибаться. Прямые мышцы бедра помогают разгибать или поднимать колено, а также сгибать бедро.Прямая мышца бедра — единственная мышца человеческого тела, которая может сгибать бедра.
C. Приводящие мышцы: приводящие мышцы состоят из четырех мышц, расположенных внутри бедра. Он связывает бедра вместе. Четыре приводящие мышцы — это длинная приводящая мышца, короткая приводящая мышца, внешняя запирательная мышца, большая приводящая мышца и тонкая мышца.
Большая приводящая мышца тянет и сокращает бедро по направлению к средней линии тела, что позволяет выполнять такие действия, как ходьба, спринт и т. Д. Длинная приводящая мышца расположена в скелете бедра.
Короткая приводящая мышца помогает сгибать и вращать бедро, сводя бедро к тазобедренному суставу. Наружная запирательная мышца вместе с другими короткими мышцами тазобедренного сустава способствует стабилизации сустава. Мышцы gracilis находятся в паху и отвечают за приведение бедра и сгибание колена.
Колено
Коленный сустав бедра и голени. Он состоит из коленных костей, коленных сухожилий и коленных связок.
Голень
Пять типов мышц включают голень, а именно икроножные, камбаловидные, подошвенные, большеберцовые и малоберцовые мышцы.
Икры являются основными мышцами икр. Gastrocnemius мышцы позволяют движение, которое помогает пальцам ног указывать вниз. Камбаловидные мышцы помогают коленям и помогают при ходьбе и стоянии.
Маленькие мышцы задней части голени называются подошвенными. Мышцы большеберцовой мышцы расположены в передней и задней части голени. В передней части голени есть только места, где расположены малоберцовые мышцы — разгибатели, задняя большеберцовая мышца, сгибатели, передняя большеберцовая и малоберцовая.
Кости ноги
Нога состоит из трех костей
Верхняя кость ноги называется бедренной костью, а нижняя часть ноги — это большеберцовая и малоберцовая кости.
Бедренная кость
Бедренная кость или бедренная кость, самая тяжелая, самая сильная и длинная во всем человеческом теле. Он может выдерживать экстремальные нагрузки, действующие на бедренную кость. Единственная кость, расположенная внутри бедра, — это бедренная кость. Бедренная кость поддерживает тазовую кость до костей голени. Самая важная особенность бедренной кости в том, что она образует шарнирное соединение с бедром.
Большеберцовая кость
Одна из двух костей голени — это большеберцовая кость. Ее еще называют голени или большеберцовой кости. Кроме того, большеберцовая кость — самая большая и крепкая кость голени. По анатомии большеберцовая кость соединяет лодыжку с коленом.
Малоберцовая кость
Малоберцовая кость, известная как кость голени, соединяется с верхней и нижней большеберцовой костью. Он расположен в боковой части ноги. Основная функция малоберцовой кости — действовать как соединение для мышц, а не как опора.Малоберцовая кость имеет три основных сочленения с латеральным мыщелком большеберцовой кости.
Набор для скелета ноги человекаAxis Scientific, анатомическая модель в натуральную величину, 36 дюймов, со всеми костями ног, съемным тазобедренным суставом и полностью шарнирной стопой, 3 года гарантии без хлопот и подробное руководство по продукту: Amazon.com: Industrial & Scientific
В настоящее время недоступен.
Мы не знаем, когда и появится ли этот товар в наличии.- Убедитесь, что это подходит введя номер вашей модели.
- МОДЕЛЬ СКЕЛЕТА НОГИ ЧЕЛОВЕКА РАЗМЕР ЖИЗНИ: Чтобы помочь вам подготовиться к следующему экзамену, презентации или демонстрации, Axis Scientific предлагает вам очень подробную копию ноги человека. Эта модель скелета человеческой ноги Axis Scientific — Right отлита из настоящей кости и имеет все естественные костные ориентиры и анатомические особенности, которые вы ожидаете найти, глядя на настоящую вещь.
- 100% АНАТОМИЧЕСКАЯ ТОЧНОСТЬ: эта модель скелета ноги человека, от бедра до пальцев ног, разработана американской командой профессионалов, чтобы обеспечить вам точную модель, на которую можно положиться в любых условиях.Как передний, так и задний вид позволяют провести полное изучение и обзор независимо от того, находитесь ли вы в классе, в полевых условиях, в офисе, в студии или дома. Куда бы вы ни пошли, вы можете положиться на эту модель кости, чтобы получить точную научную информацию.
- ВЗАИМОДЕЙСТВУЙТЕ И ИЗУЧАЙТЕСЬ С ПРОДУКТАМИ, СОЗДАННЫМИ ДОЛГОВЕЧНЫМИ: Хотя каждое человеческое тело уникально по-своему, этот скелет Axis Scientific Leg — именно то, что вам нужно, чтобы начать понимать человеческое тело более глубоко.Эта костяная модель выполнена из прочного пластика на основе ПВХ, поэтому ее можно брать с собой куда угодно, не беспокоясь о ее прочности и долговечности.
- ГОТОВЫ ВЫЙТИ ИЗ КОРОБКИ: Хотя этот скелет ноги может показаться простым инструментом анатомического исследования, мы всегда хотим, чтобы вы были готовы. Каждая модель Axis Scientific разработана так, чтобы быть простой и доступной для новичков, при этом сохраняя высокое качество и для профессионалов.Банджи и проволочная фурнитура этой модели обеспечивают полную артикуляцию и подробное изучение.
- 3-ЛЕТНЯЯ ГАРАНТИЯ И ПОДДЕРЖКА, НА КОТОРЫЕ МОЖНО РАСЧЕТАТЬ: Axis Scientific с гордостью представляет свою 3-летнюю гарантию без проблем, но наша служба поддержки клиентов не останавливается на достигнутом. Мы ценим наших клиентов, и наша специальная команда поддержки готова помочь им с любой потребностью столько, сколько они вам понадобятся. В отличие от многих других компаний, мы базируемся в Соединенных Штатах и позаботимся о том, чтобы вы никогда не остались без модели, благодаря нашей непрерывной программе ремонта или замены.
Характеристики этого продукта
Фирменное наименование Axis Scientific Цвет Костно-белый Ean 0616316879508 Вес изделия 4.45 фунтов Номер модели A-105181 Кол-во позиций 1 Номер детали A-105181 Размер В натуральную величину (длина 36 дюймов) Код КПСС ООН 41000000 UPC 616316879508 Что следует знать перед протезированием ноги
Физиотерапия Реабилитационная терапияОтзыв от:
Протезы ног или протезы могут помочь людям с ногами. ампутации легче передвигаться.Они имитируют функцию, а иногда даже появление настоящей ноги. Некоторым людям все еще нужна трость, ходунки или костыли. ходить с протезом ноги, а другие могут ходить свободно.
Если вам предстоит ампутация нижней конечности или она скоро произойдет, протез ноги вероятно, вариант, о котором вы думаете. Специалист по реабилитации ампутантов Мэри Кеслер, доктор медицины, делится несколькими соображениями, которые следует учесть в первую очередь.
Не всем выгоден протез ноги
Хотя многие люди с потерей конечностей хорошо себя чувствуют с протезами ног, каждый — хороший кандидат на протез ноги.Несколько вопросов вы можете хотите обсудить с врачом, прежде чем выбирать протез ноги включают:
- Достаточно ли мягкой ткани, чтобы смягчить оставшуюся кость?
- Сколько тебе боли?
- Какое состояние кожи на конечности?
- Каков диапазон движений остаточной конечности?
- Другая нога здорова?
- Каким был уровень вашей активности до ампутации?
- Каковы ваши цели мобильности?
Тип ампутации (выше или ниже колена) также может повлиять на ваше решение.Обычно проще использовать протез ноги ниже колена, чем протез выше колена. «Если коленный сустав не поврежден, протез нога требует гораздо меньше усилий для движения и обеспечивает большую подвижность », — поясняет Кеслер.
Причина ампутации также является фактором, поскольку она может повлиять на здоровье остаточной конечности. Ваше физическое здоровье и образ жизни также важно учитывать. Если вы были не очень активны и потеряли ногу из-за заболевание периферических сосудов или диабет, например, вы будете бороться с протезом больше, чем тот, кто был очень активен, но потерял конечность в автокатастрофе.
Что касается ампутации, каждый человек уникален. Решение переехать вперед с протезом должен быть совместным между вами и ваш доктор.
Чтобы получить правильный тип и подгонку, важно работать в тесном сотрудничестве со своим протезистом — отношения, которые могут быть у вас на всю жизнь.
Мэри Кеслер, доктор медициныПротезы ног не подходят для всех
Если ваш врач пропишет протез ноги, вы можете не знать, где его начинать.Это помогает понять, как работают разные части протеза. вместе:
- Сам протез ноги изготовлен из легких, но прочных материалов. В зависимости от местоположения ампутации, нога может иметь или не иметь функциональное колено и голеностопные суставы.
- Розетка представляет собой точный слепок вашей остаточной конечности, который плотно прилегает к конечности. Это помогает прикрепить протез ноги к вашему телу.
- Система подвески как протез остается прикрепленным, будь то всасывание рукава, вакуумная подвеска / отсасывание или дистальная блокировка с помощью штифта или шнурка.
Существует множество вариантов для каждого из вышеперечисленных компонентов, каждый из которых свои плюсы и минусы. «Чтобы подобрать нужный тип и размер, важно работать в тесном контакте со своим протезистом — отношения, которые у вас могут быть жизнь », — рекомендует Кеслер.
А протезист специалист в области здравоохранения, специализирующийся на протезах конечностей и способный помочь вам выбрать правильные компоненты. У вас будут частые встречи, особенно вначале, поэтому важно чувствовать себя комфортно с протезист выбираете вы.
Реабилитация — это непрерывный совместный процесс
После того, как вы выбрали компоненты протеза ноги, вам понадобится реабилитация для укрепления ног, рук и сердечно-сосудистой системы, а также вы учитесь ходить с новой конечностью. Вы будете тесно сотрудничать с врачи-реабилитологи, физиотерапевты и эрготерапевты разработать план реабилитации с учетом ваших целей мобильности. Большая часть этого плана — поддерживать здоровую ногу в тонусе. «Твоя здоровая нога на вес золота », — подчеркивает Кеслер.«Пока протез технологии постоянно развиваются, ничто не может воспроизвести здоровую ногу ».
Комплексная программа реабилитации инвалидов Джонса Хопкинса
При восстановлении после ампутации конечности необходима поддержка специальной группы экспертов. В Johns Hopkins наша команда физиологов, ортопедов, протезистов, физиотерапевтов и эрготерапевтов, психологов-реабилитологов и других специалистов работает вместе над созданием индивидуального плана реабилитации.
Привыкнуть к протезу ноги непросто
Научиться пользоваться протезом ноги может быть непросто. Даже после первоначальная реабилитация окончена, вы можете столкнуться с некоторыми проблемами, которые Команда протезистов и реабилитологов поможет вам справиться. Общие препятствия включают:
- Чрезмерное потоотделение (гипергидроз), который может повлиять на посадку протеза и привести к проблемам с кожей.
- Изменение формы остаточной конечности . Обычно это происходит в первый год после ампутации. ткань принимает более постоянную форму и может повлиять на прилегание розетка.
- Слабость в остаточной конечности, что может затруднить использование протеза в течение длительного периода время.
- Фантомная боль в конечностях может быть достаточно интенсивным, чтобы повлиять на вашу способность пользоваться протезом.
Заметка о фантомной боли в конечностях
Фантомная боль в конечности или боль, которая, кажется, исходит от ампутированной конечности, — это очень реальная проблема, с которой вы можете столкнуться после ампутации. «Около 80% люди с ампутациями испытывают фантомную боль в конечностях, не имеющую явного причиной, хотя боль в конечности перед ампутацией может быть фактором риска », — говорит Кеслер.
Зеркальная терапия , при которой вы выполняете упражнения с зеркалом, может помочь при определенных типах фантомная боль в конечностях.«Взгляд на себя в зеркало имитирует наличие ампутированной ноги, заставляющее мозг думать, что она все еще там, останавливая боль », — объясняет Кеслер.
В других случаях фантомная боль в конечностях может быть вызвана другим заболеванием. поражает остаточную конечность, например радикулит или неврома. Устранение этих первопричин может помочь устранить фантом боль.
Ваши потребности в протезе ноги могут измениться
В какой-то момент вы можете заметить, что не так функциональны, как хотелось бы быть с вашим текущим протезом ноги.Может быть, у вашей остаточной конечности стабилизируется, и вы готовы перейти от временного протеза, который длится от нескольких месяцев до одного, который может длиться от трех до пяти лет. Или, может быть вы «обошли» свой протез, двигаясь больше или иначе, чем протез предназначен для. Новая боль, дискомфорт и нестабильность некоторые признаки того, что, возможно, пора обратиться к протезисту, чтобы переоценить свои потребности.
Ваш протезист может порекомендовать отрегулировать ваше текущее оборудование или замена одного из компонентов.Или вы можете получить рецепт на новый протезирование ноги, которое происходит в среднем каждые три-пять лет. если ты получать новые компоненты, важно найти время, чтобы понять, как они работают. Физическая терапия может помочь приспособиться к новым компонентам или вашему новый протез ноги.
Технологии протезирования ног постоянно развиваются
Всегда есть новые разработки в технологии протезирования конечностей, такие как компоненты, управляемые микропроцессором, и компоненты, ориентированные на конкретную деятельность.
- Муфты микропроцессорные оснащены компьютерными микросхемами и датчиками, чтобы походка была более естественной.Они могут даже иметь разные режимы для ходьбы по плоской поверхности или вверх и вниз по лестнице.
- Существуют также специализированные протезы ног для различных видов деятельности , таких как бег, душ или плавание, на которые можно переключаться по мере необходимости. В некоторых случаях, Ваш повседневный протез ноги может быть изменен вашим протезистом для обслуживания разные цели.
- Операция остеоинтеграции это еще один вариант. Эта процедура предполагает введение металлического имплантат прямо в кость, поэтому нет необходимости в гнезде.В Затем протез ноги прикрепляется непосредственно к этому имплантату. Пока это процедура подходит не всем и все еще изучается, она может обеспечивают улучшенный диапазон движений и сенсорное восприятие.
Важно помнить, что вы не одиноки в навигации по многим различные варианты протезирования ног. Ваша группа по уходу поможет вам взвесить плюсы и минусы каждого и выберите идеальный протез ноги, который подходит ваш образ жизни.
IJERPH | Бесплатный полнотекстовый | Длина ног, пропорции тела и здоровье: обзор с заметкой о красоте
1.Введение
Бесстрастный натуралист с другой планеты, выполняющий миссию по сбору на Землю, может быть удовлетворен одним или двумя образцами Homo sapiens как представителя этого вида. Человеческие наблюдатели нашего вида не так-то легко успокаиваются. Это связано с тем, что с антропоцентрической точки зрения люди обладают разнообразными размерами, формами, цветами, темпераментом и другими фенотипическими характеристиками. Профессиональные антропологи, врачи и другие специалисты веками обсуждали причину и значение фенотипической изменчивости человека.Большая часть исторического дискурса была сосредоточена на концепциях «расы», а часть споров была сосредоточена на человеческом статусе различных живых групп людей [1,2]. Серьезные предложения относительно иерархии человечности появились совсем недавно, в 1962 году, с публикацией «Происхождения рас» Карлтона Куна [3], профессора антропологии Пенсильванского университета. Кун разделил живые народы мира на пять «рас», в частности, по размеру и пропорциям тела. Австралийские аборигены (называемые кунами «австралоиды») имеют исключительно длинные ноги по сравнению с ростом, а африканские пигмеи («конгоиды» в таксономии кунов) имеют исключительно низкий рост, длинные руки относительно длины ног и особенно короткие голени. .По словам Куна, «их манера затмевать грани ахондропластики…» [3, с. 653]. Более того, Кун утверждал, что обе «расы» пересекли порог видов между Homo erectus и H. sapiens только за последние 10 000–50 000 лет. Кун, напротив, предположил, что древние европейцы (которых Кун назвал «европеоидами») переступили порог H. sapiens около 200 000 лет назад. По словам Куна, древние европейцы были «нормальными» по размеру и форме, могли «… сидеть в любом западноевропейском ресторане, не вызывая особых комментариев, за исключением их манер за столом» [3, с.582]. Эти претензии на расовую таксономию человека, включая временные пороги Куна для гомо-сапиенации, были дискредитированы палеонтологическими и геномными исследованиями, показывающими древность происхождения современного человека в Африке, а также существенную геномную африканскую природу всего живого. люди [4–6]. Утверждение Куна о том, что африканские пигмеи имеют «ахондропластические пропорции», также неверно. Ши и Бейли [7] показывают, что африканские пигмеи уменьшены в общем размере и имеют форму тела, которая аллометрически пропорциональна уменьшению размера.Отказ от расистской истории изучения морфологии человека позволил исследованию сосредоточиться на более значимых биологических, медицинских, социальных и эстетических последствиях размера и формы человеческого тела. В этой статье мы рассматриваем доказательства того, что форма человеческого тела, особенно длина ног по отношению к общему росту, является важным показателем для эпидемиологии и экологического здоровья населения. Мы обнаружили, что у всех людей, а также в пределах географических, социальных и этнических групп людей относительная длина ног отражает состояние питания и здоровье в годы физического роста, а также имеет биологически и статистически значимые связи с рисками заболеваемости и смертности. в зрелом возрасте.
4. История эволюции формы человеческого тела
Человеческий вид отличается от нечеловеческих приматов несколькими анатомическими особенностями. Среди них пропорции рук и ног по отношению к общей длине тела. Различия между людьми проиллюстрированы на рисунке 5. Пропорционально общей длине тела, измеряемой как рост, современные взрослые люди имеют относительно длинные ноги и короткие руки. Количественные различия между взрослыми людьми, шимпанзе (Pan troglodytes) и бонобо (Pan paniscus) приведены в таблице 1.Комбинированные значения интермембрального индекса и плечево-бедренного индекса показывают, что у людей кости ног в среднем на 34% длиннее, чем у нечеловеческих обезьян, относительно длины костей рук. Основная причина этого — передвижение человека на двух ногах, поведение, которое развилось по крайней мере 4,4 миллиона лет назад (MYA), как показано на ископаемом виде гоминина Ardipithecus ramidus. Длина ноги должна составлять примерно 50 процентов от общего роста, чтобы достичь биомеханической эффективности шагающей двуногой походки человека.У современных людей это происходит в конце стадии истории жизни детства, которая наступает примерно в 7,0 лет [11]. К зрелому возрасту человеческие видоспецифичные пропорции тела позволяют не только ходить на двух ногах, но также, как это было замечено, экспериментально проверено или предположительно предполагалось, для технологических манипуляций [12], более эффективной терморегуляции в условиях тропической саванны [13]. –16], освобождение рук для переноски предметов и младенцев [17], для бега на длинные дистанции [18], а также для жестикуляции, общения, языка и социально-эмоционального контакта [19].Пропорции тела взрослого человека обусловлены разным ростом сегментов тела [21]. При рождении длина головы составляет примерно одну четверть от общей длины тела, в то время как в возрасте 25 лет длина головы составляет только примерно одну восьмую общей длины. Также пропорционально изменяется длина конечностей, которые становятся длиннее по отношению к общей длине тела в годы роста [22]. На рисунках на Рисунке 6 показаны типичные изменения, которые происходят у людей от рождения до 25 лет.Люди следуют цефалокаудальному градиенту роста и развития, типичному для большинства млекопитающих. Однако есть некоторые видоспецифические особенности развития плана человеческого тела. В классической статье 1926 года Шульц [23] опубликовал свои наброски пропорций тела зародышей гоминоидов, воспроизведенные здесь как рисунок 7. Человеческий плод «4 -го месяца» имеет относительно более короткие ноги, чем шимпанзе, орангутан или гиббон. . Это предполагает, что оценки Шульца развития обезьян, не являющихся людьми, верны (см. Легенду на Рисунке 7).Другое отличие, не отмеченное Шульцем, — это пропорциональный размер черепа относительно лица, который у человеческого плода больше, чем у шимпанзе, орангутана или гиббона. затем к зрелости можно частично объяснить эволюцией двуногости, взаимодействующей с эволюцией большого и сложного мозга. У обезьян характерен быстрый рост мозга до рождения и относительно медленный после рождения. У людей быстрый рост мозга как до, так и после рождения [24,25].У новорожденных людей мозг больше, чем у обезьян, хотя и не намного больше с точки зрения соотношения массы мозга и тела (таблица 2). Различия между человеком и обезьяной в массе мозга и соотношении массы мозга к массе тела намного больше во взрослом возрасте, и большая часть этих различий фактически достигается к 6,9 годам [11,25]. Пожалуй, решающее различие заключается не только в массе мозга, но и в его метаболической активности. Новорожденный человек использует 87% своей скорости метаболизма в состоянии покоя (RMR) для роста и функционирования мозга. К 5 годам процент использования RMR все еще высок и составляет 44%, тогда как у взрослого человека этот показатель составляет от 20 до 25% от RMR.На сопоставимых стадиях развития значения RMR для шимпанзе составляют около 45, 20 и 9% соответственно [26]. С такими высокими метаболическими потребностями мозга человеческий младенец и ребенок вполне могли быть выбраны естественным образом, чтобы пойти на компромисс в распределении ограниченных питательных веществ, кислорода и других ресурсов, необходимых для роста мозга, по сравнению с другими частями тела. Компромиссы между ростом, развитием и созреванием частей тела характерны для всего разнообразия историй жизни животных и растений [27–29], включая человеческий вид [30,31].С этой точки зрения основная причина задержки роста человеческих ног во время внутриутробного и младенческого развития заключается в том, что это способствует быстрому росту мозга. известный. Вероятно, что здесь задействованы генетические, гормональные и питательные факторы. В обзоре биологии роста костей Раух [32, с. 194] утверждает: «Рост кости в длину в основном достигается за счет действия хондроцитов в пролиферативной и гипертрофической зонах пластинки роста.Продольный рост контролируется системными, местными паракринными и местными механическими факторами. Что касается последнего, должен существовать механизм обратной связи, который гарантирует, что рост кости идет в направлении преобладающих механических сил. Как это работает, в настоящее время неизвестно ». Известно, что длина пролиферативных столбов в пластине роста коррелирует с длиной конечностей: «… вид с длинными ногами и короткими руками имеет более длинные столбики в коленях и короче в локтях, чем виды с противоположными пропорциями» [33 , п.21]. Количественное картирование локуса признаков (QTL) лабораторных мышей выявило области генома, связанные с фенотипическими различиями в длине бедренной кости, голени, плечевой кости и локтевой кости [34]. Известно, что изменения в регуляции роста генома, такие как паттерны экспрессии Hox, связаны с ростом сегментов предплечий приматов [35]. Изменения чувствительности пластинок роста костей к факторам, стимулирующим и ингибирующим рост, в разное время во время развития и в разных участках скелета, также, как известно, ответственны за дифференцированный рост сегментов тела [36,37].Еще одно предположение состоит в том, что кровообращение плода может способствовать компромиссу между ростом мозга и ног. Кровь в восходящей аорте плода имеет более высокую насыщенность кислородом, чем кровь, спускающаяся в общую подвздошную артерию (рис. 8). Кроме того, пупочные артерии несут часть крови, спускающейся к ноге, обратно к плаценте. Этот паттерн кровообращения плода характерен для большинства млекопитающих и, вероятно, является эволюционно древним. В сочетании с недавно сформировавшимися метаболическими потребностями головного мозга человеческого плода, древний паттерн кровообращения может оставлять ноги человеческого плода с пониженным поступлением кислорода и питательных веществ, что еще больше замедляет рост и развитие ног по сравнению с более крупными областями тела.Мы не можем найти экспериментальной поддержки этого предложения. Имеются данные клинического исследования на людях, свидетельствующие о том, что усиление кровотока в конечностях связано с большим ростом [38].5. Размер и форма у живых людей
Общая модель развития формы человеческого тела — это видоспецифическая характеристика. Исторические произведения искусства, скульптуры и анатомические рисунки из Европы эпохи Возрождения [40,41] и доколумбовой Мексики [42] демонстрируют фундаментальные общие черты в изображении формы тела поздних плодов, новорожденных и младенцев.Однако отдельные популяции живых людей отличаются разнообразием размеров и форм тела. Средний рост для популяций взрослых варьируется от минимальных значений для африканских пигмеев Эфе (144,9 см для мужчин и 136,1 см для женщин [43]) до максимальных значений для голландцев в Европе, составляющих 184,0 см для мужчин и 170,6 см для женщин [44]. ]. Существуют также биологически и статистически значимые различия в форме тела между популяциями людей. Эвелет и Таннер [45,46] опубликовали данные о пропорциях тела и длине ног, рассчитанные с помощью соотношения роста сидя, по десяткам человеческих популяций, распределенных по большинству географических регионов мира (рис. 9).Отношение высоты сидения (SHR) — широко используемый показатель пропорции тела. Измеренный рост минус рост в сидячем положении также можно использовать для оценки длины ног, но этот показатель не стандартизирован для общего роста, что затрудняет сравнение людей с разным ростом. Среднее значение SHR для популяций взрослых варьируется от минимальных значений, т. Е. Относительно самых длинных ног для австралийских аборигенов (SHR = 47,3 для мужчин и 48,1 для женщин), до максимальных значений SHR, т. Е. Относительно самых коротких ног, для мужчин майя из Гватемалы и перуанских женщин. (SHR = 54.6 и 55.8). Осмысление этих сравнений во всем мире затруднено из-за различий в образе жизни, окружающей среде и геномике. Два хорошо известных экогеографических принципа, правила Бергмана и Аллена, часто упоминаются как основные причины глобальных закономерностей изменения формы человеческого тела. Бергманн [47] в 1847 году заметил, что близкородственные виды млекопитающих, такие как медведи, имеют большую массу тела в более холодном климате. Аллен [48] добавил в 1877 году, что конечности и хвосты таких видов обычно короче в холодном климате и длиннее в более теплых.Большая масса тела и относительно короткие конечности увеличивают соотношение объема к площади поверхности и обеспечивают форму тела, которая максимизирует метаболическое удержание тепла у млекопитающего. И наоборот, при более высоких температурах относительно длинные конечности увеличивают площадь поверхности по сравнению с объемом и допускают большие потери тепла. Экспериментально было показано, что мыши и другие млекопитающие, кроме человека, выращенные в более высоких температурах, испытывают больший рост костной ткани и более длинные кости конечностей [49]. Обычное объяснение этого — большая васкуляризация, обеспечивающая большую перфузию кислорода и питательных веществ.Однако недавние экспериментальные исследования показывают, что даже при отсутствии сосудистой сети, культура хондроцитов плюсневой кости мыши in vitro демонстрирует положительную корреляцию между температурой окружающей среды и «… большей пролиферацией и объемом внеклеточного матрикса…» [49, с. 19348]. Правила Бергмана и Аллена в некоторой степени применимы к человеческому виду. В 1953 году Робертс [50] опубликовал анализ, показывающий значительную взаимосвязь между массой тела и широтой для людей, с группами людей, живущих в более высоких широтах, имеющих большую массу тела, чем у людей, живущих ближе к экватору.Двадцать пять лет спустя Робертс [51] обновил и подтвердил эти результаты. Другое исследование показывает, что люди, живущие в более холодных регионах, также имеют более короткие конечности относительно общего роста по сравнению с группами людей, живущих в более теплых регионах [15,45,46]. Однако эти климатические отношения не идеальны. Повторный анализ данных Робертса, проведенный Кацмарзиком и Леонардом [52], изменяет важность климата как основного фактора, определяющего форму человеческого тела. Кацмарзик и Леонард анализируют соотношение высоты сидящего в 165 группах людей, изученных в период с 1960 по 1996 год.Все данные о людях, проанализированные Робертсом, были собраны до 1953 года. Кацмарзик и Леонард показывают, что недавно изученные группы по-прежнему следуют экологическим принципам формы тела, но что связь с климатом ослабла после исследования Роберта. Наклон наиболее подходящих линий линейной регрессии для отношения средней годовой температуры к соотношению высоты сидящего вдвое меньше, чем у Робертса. Кацмарзик и Леонард (стр. 483) утверждают, что «… хотя климатические факторы продолжают оставаться значительными корреляциями мировых вариаций размеров и морфологии человеческого тела, различия в питании среди тропических популяций развивающихся стран уменьшили их влияние.Авторы определяют изменения в питании как изменения в диете и образе жизни, особенно введение западных продуктов питания и поведения. Они отмечают, что «… климат может формировать морфологию, влияя на доступность продуктов питания и питание, [что означает, что] линейное строение тропических популяций является следствием [факторов] питания, а не теплового стресса…» (стр. 491–492). В этом случае за годы роста и развития более или менее общее потребление пищи, большее или меньшее количество любых необходимых питательных веществ, большая или меньшая физическая активность (и тип активности) могут повлиять на форму тела.Гватемала Майя, например, потребляют только около 80% всей энергии, необходимой для здорового роста, а 20,4% также испытывают дефицит йода [53]. Дефицит йода в младенчестве и детстве приводит к уменьшению длины ног, особенно дистального отдела бедра, голени и стопы [54]. Дети и взрослые майя тратят много времени и энергии на тяжелый труд [55], который отвлекает доступную энергию в рационе от роста. Известно, что такое сочетание питания и образа жизни снижает общий рост и длину ног [56].Форма тела людей может иметь генетическую основу, особенно для групп людей, которые проживали в одной среде на протяжении многих поколений. Сравнение роста и пропорций тела между чернокожими (афроамериканцы) и белыми (американцы европейского происхождения) в Соединенных Штатах дает пример взаимодействия генома и окружающей среды и их влияние на рост [57]. Опубликованные данные первого Национального обследования здоровья и питания (NHANES I) в Соединенных Штатах собрали антропометрические данные о репрезентативной на национальном уровне выборке чернокожих и белых в возрасте от 18 до 74 лет.Когда данные скорректированы с учетом различий между двумя этническими группами по доходу, образованию, проживанию в городе или деревне и возрасту, не будет значительной разницы в среднем росте между чернокожими и белыми мужчинами. Также нет существенной разницы в среднем росте между черными и белыми женщинами. Хотя белые и черные взрослые в Соединенных Штатах имеют одинаковый средний рост, при контроле образования, дохода и других переменных пропорции тела в этих двух группах различаются. Крогман [58] обнаружил, что при одинаковом росте у чернокожих, живущих в Филадельфии, США, туловище и конечности короче, чем у белых, особенно голени и предплечья.Hamill et al. [59] обнаружили, что это верно и для национальной выборки чернокожих и белых молодых людей в возрасте от 12 до 17 лет, а также для взрослых в возрасте 20-49 лет, измеренных в обзоре NHANES III, 1988–1994 гг. [9]. Геномный вклад в различия пропорций тела между черными и белыми кажется вероятным, поскольку чернокожие, как правило, имеют большее геномное происхождение из Африки к югу от Сахары, чем белые. При статистическом анализе родословных двух образцов человека Лившиц и соавт.[60] подсчитали, что от 40% до 75% межличностных вариаций пропорций тела, которые они изучали (с поправкой на возраст и пол), объясняются «генетическими эффектами». Это может быть лучше описано как семейные эффекты, потому что авторы проанализировали семьи, а также потому, что они обнаружили значительные общие экологические эффекты для братьев и сестер, а также значимые зависимости пола от возраста. Диапазон источников вариабельности в анализе затрудняет вычисление простой генетической вариативности. Даже если обнаруживаются конкретные генотипы, их прямой вклад в нормальные этнические (так называемые «расовые») вариации формы человеческого тела может быть относительно небольшим.На 40 неделе беременности плоды, идентифицированные как афроамериканцы, в среднем имеют относительно более длинные ноги, чем плоды, идентифицированные как американцы европейского происхождения [23]. Но разница, измеряемая по (общая длина / длина макушки), составляет менее 1%. При анализе данных, показанных на рисунке 9, Bogin et al. [61] оценили вклад географического происхождения в дисперсию SHR в 0,04, что хорошо согласуется с геномными оценками вариации в общем росте 0,04–0,06 [2]. Судебные антропологи и врачи в Соединенных Штатах часто использовали «расовые» пропорции тела, чтобы приписать скелету афроамериканскую или европейскую / азиатско-американскую этническую принадлежность [63,64].Feldesman and Fountain [65] проверили полезность соотношения длина / рост бедренной кости, чтобы правильно идентифицировать 798 пар бедренная кость / рост скелетов известной этнической принадлежности. Они обнаружили, что «… соотношение бедренной кости к росту у« черных »статистически значимо отличается от такового у« белых »и« азиатов »[стр. 207]. Дискриминантная функция и кластерный анализ показывают, однако, что согласованность групп, определенных географическим происхождением, невысока, а результаты едва ли лучше случайности. Использование «расовых» пропорций тела для идентификации неизвестных скелетов привело бы к большому количеству неверных указаний этнической принадлежности.Более многообещающий подход к пониманию контроля пропорций человеческого тела исходит из геномных исследований. Hox-гены и гомеобоксовые последовательности, а также растущее число факторов роста и передачи сигналов, как известно, регулируют рост сегментов тела [66], и эти гены являются общими для всех таксонов. Имеются наблюдательные и экспериментальные доказательства того, что экспрессия Hoxd связана с различиями в длине предплечья, кисти и пальцев у обезьян [35]. Ген, содержащий гомеобокс низкорослого роста (SHOX), является еще одной областью генома, которая может иметь отношение к пропорциям человеческого тела.«SHOX, расположенный на дистальных концах хромосом X и Y, кодирует фактор транскрипции гомеодомена, ответственный за значительную часть роста длинных костей [67]. Синдром Тернера (45, кариотип XO) приводит к дефициту роста примерно на 20 см. Некоторые исследования показывают, что ноги страдают непропорционально [68,69], но другие исследования не обнаруживают диспропорции [70]. Более конкретные гены-кандидаты, определяющие форму тела, известны у некоторых млекопитающих, кроме человека [71,72], и у насекомых [27]. Другой очень активной областью исследований является эпигенетическая регуляция роста тела [73].Эффекты эпигенома могут действовать через ряд взаимодействий генома (например, метилирование ДНК и модификация гистонов), протеома (например, регуляция экспрессии генов микро-РНК) и окружающей среды (например, климат, диета и физическая активность) и вполне могут играть роль важную роль в определении размера и формы человека.6. Пластичность развития
Пластичность относится к концепции, согласно которой развитие фенотипа организма зависит от изменений качества и количества факторов окружающей среды, необходимых для жизни [74].Мы используем эту концепцию здесь, чтобы обозначить, что за годы роста и развития у человека может вырасти больше или меньше различных тканей, и они станут взрослыми разных размеров и форм. У взрослых эти размеры и формы в основном фиксированы, особенно в отношении общего роста и длины сегментов тела. Человеческий рост очень пластичен в годы роста и развития, что соответствует общему качеству условий жизни [11]. С точки зрения пластичности развития, длина ног, как с точки зрения абсолютного размера, так и относительно общего роста, является индикатором качества среды для роста в младенчестве, детстве и юношеском возрасте.Причиной этого является общий принцип, согласно которому те части тела, которые растут быстрее всего, больше всего пострадают от нехватки питательных веществ, инфекции, паразитов, физических или эмоциональных травм и других неблагоприятных условий. Цефало-каудальный принцип роста применительно к человеческому виду означает, что ноги, особенно большеберцовые кости, растут быстрее по сравнению с другими сегментами тела от рождения до возраста 7 лет. Таким образом, относительно короткий LL у подростков и взрослых может быть вызван неблагоприятными условиями в младенчестве и детстве, ведущими к конкуренции между сегментами тела, такими как туловище и конечности, а также между органами и конечностями.В простейшем случае такая конкуренция может быть за ограниченные питательные вещества, доступные во время роста [31,56,61]. Более сложные объяснения конкуренции относятся к аспектам гипотезы бережливого фенотипа [75,76], гипотезы межпоколенческих влияний [77,78], гипотезы программирования плода [79] и гипотезы прогнозирующего адаптивного ответа [80,81]. Обсуждение этих гипотез выходит за рамки данного обзора [см. Ссылку 31 и другие статьи в том же выпуске для такого обсуждения], но по сути каждая из этих гипотез предсказывает, что жизненно важные органы головы, грудной клетки и брюшной полости тело будет защищено от невзгод за счет менее жизненно важных тканей конечностей.8. Длина ног и красота
«Ноги, помимо того, что являются очень важной функциональной единицей, сами по себе являются важным сексуальным влечением, и во всех культурах они занимают преобладающее место в концепции красоты» [134, с. 505]. Забота о пропорциях тела имеет глубокие корни в европейской истории. Основываясь на работах Витрувия в первом веке до нашей эры. Римский архитектор и писатель Леонардо да Винчи (1452–1519) разработал каноны или правила рисования человеческих пропорций. Согласно этим канонам, рост человека должен составлять восемь голов с дополнительной четвертью головы на длину шеи.Длина ног должна составлять четыре длины головы. «Витрувианский человек» Леонардо (ок. 1487 г.) является знаковой иллюстрацией канонов. Альбрехт Дюрер (1471–1528), немецкий художник, разработал технологию рисования как канонических форм, так и множества вариаций, наблюдаемых в природе. С помощью своих геометрических методов Дюрер мог нарисовать любые человеческие вариации в размерах и пропорциях. Он применил свой метод к рисункам мужчин, женщин, детей и младенцев. Включение женщин и детей в этот тип методологической работы было новшеством, поскольку большинство художников следовали учению Ченнино Ченнини (ок.b1400), который писал, что женщины «… не имеют определенной пропорции» [40, с. 202]. Похоже, дети были слишком незначительны, чтобы Ченнини даже упомянул!После 1600 года художники постренессанса начинают изображать детей с нормальными пропорциями, а также с патологиями роста. Фламандский художник Ван Дайк изображает троих нормальных детей на картине «Дети Карла I» (1635 г.). На картине Диего Веласкеса «Фрейлины» (1656 г.) изображены нормальный ребенок, женщина с ахондропластической карликовостью (голова и туловище нормальных размеров с короткими руками и ногами) и мужчина с карликовостью, вызванной дефицитом гормона роста (пропорциональное уменьшение в размерах). всех частей тела).Во время этих картин, конечно, не было известно о биологическом контроле нормального и патологического роста в размерах и пропорциях.
Эдмунд Берк, британский государственный деятель и философ, опубликовал в 1756 году эссе «Философское исследование происхождения наших представлений о возвышенном и прекрасном». Одна часть этого эссе имеет подзаголовок «Пропорция, а не причина красоты у человека». Берк утверждал, что людей с пропорциями тела, выходящими за рамки канона Леонардо, все еще можно считать красивыми.Он считал человеческую ногу особенно красивой: «Я полагаю, что никто не подумает, что форма ноги человека так хорошо приспособлена для бега, как ноги лошади, собаки, оленя и некоторых других существ; по крайней мере, у них не такой внешний вид: все же, я полагаю, хорошо сделанная человеческая нога будет позволена намного превосходить все это по красоте ». Остается задаться вопросом, какие человеческие ноги «хорошо вылеплены». Возможно, Берк имел в виду относительно прямые и длинные — противопоказания к рахиту, предполагающие хорошее здоровье и хорошее питание в детстве и предсказывающие плодовитость у взрослых женщин.
Пересечение биомедицинских и эстетических проблем с красотой человеческой ноги по-прежнему остается сильным. Цитата Cuenca-Guerra и его коллег [134], открывающая этот раздел, взята из статьи о хирургическом использовании имплантатов голени для повышения привлекательности ног. Растет количество литературы по научному анализу красоты и медицинских средств ее улучшения, большая часть из которых посвящена пропорциям тела и длине ног (например, [135–137]).9. Заключение
Косметическая хирургия, высокие каблуки на обуви и другие умные стили одежды могут сделать ноги более привлекательными, но эти методы не преодолевают фундаментальную связь между длиной ног и здоровьем человека.Обширный обзор литературы показывает, что существуют убедительные доказательства того, что взрослые с диспропорциями скелета, особенно с высоким SHR (короткие ноги), подвергаются большему риску ишемической болезни сердца (ИБС) из-за гиперхолестеринемии, нарушения регуляции уровня глюкозы и инсулина, повышения пульсового давления и систолическое артериальное давление и более высокий уровень фибриногена [103]. Некоторые виды рака связаны с относительно длинными ногами. Пренатальное и послеродовое недоедание и болезни являются причиной относительно коротких ног у взрослых, но все еще не объясняют, почему они подвергаются большему риску заболеваний и смертности в более раннем возрасте, чем взрослые с более длинными ногами.Связь между задержкой роста в детстве и избыточной массой тела у взрослых становится хорошо известной. Проспективное трехлетнее исследование низкорослых бразильских мальчиков и девочек в возрасте 11–15 лет показало, что они набирают больше жировой массы и меньше мышечной массы по сравнению со сверстниками без задержки роста [138]. Взрослые бразильские женщины с низким ростом и непропорционально короткими ногами имеют высокий риск ожирения [139]. Причина такой связи с ожирением, по-видимому, связана с нарушением окисления жиров у детей с задержкой роста [140]. Дыхательный коэффициент натощак (RQ = отношение объема углекислого газа, производимого организмом к объему потребляемого кислорода) значительно выше, и, следовательно, окисление жиров ниже, что приводит к большим запасам жира в организме в группе низкорослых.